

AGG/CCT interruptions affect nucleosome formation and positioning of healthy-length CGG/CCG triplet repeats
- PMID: 24261641
- PMCID: PMC3870987
- DOI: 10.1186/1471-2091-14-33
AGG/CCT interruptions affect nucleosome formation and positioning of healthy-length CGG/CCG triplet repeats
Abstract
Background: Fragile X Syndrome (FXS), the most common inherited form of mental retardation, is caused by expansion of a CGG/CCG repeat tract in the 5'-untranslated region of the fragile X mental retardation (FMR1) gene, which changes the functional organization of the gene from euchromatin to heterochromatin. Interestingly, healthy-length repeat tracts possess AGG/CCT interruptions every 9-10 repeats, and clinical data shows that loss of these interruptions is linked to expansion of the repeat tract to disease-length. Thus, it is important to understand how these interruptions alter the behavior of the repeat tract in the packaged gene.
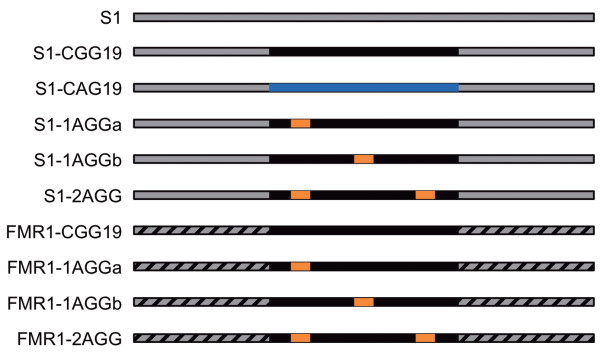
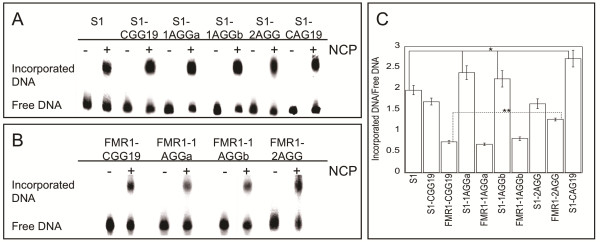
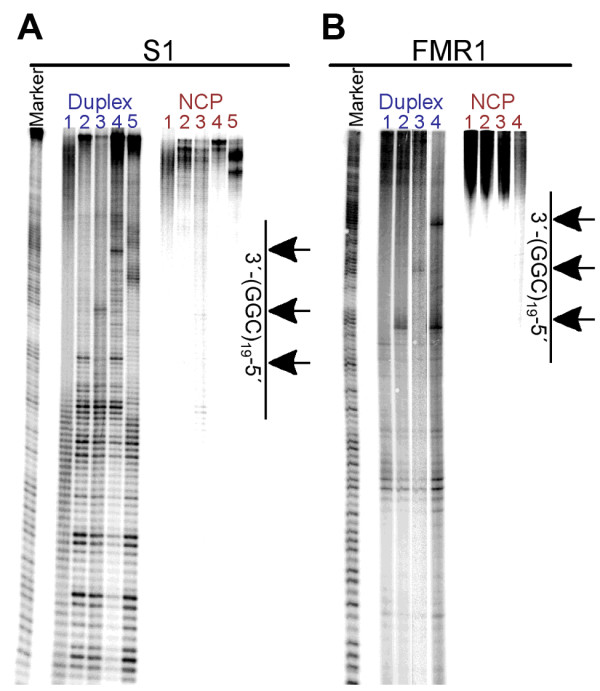
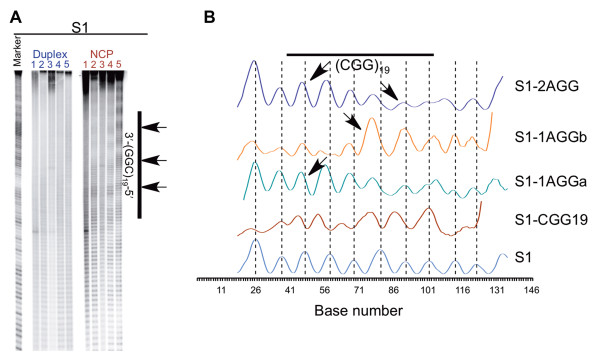
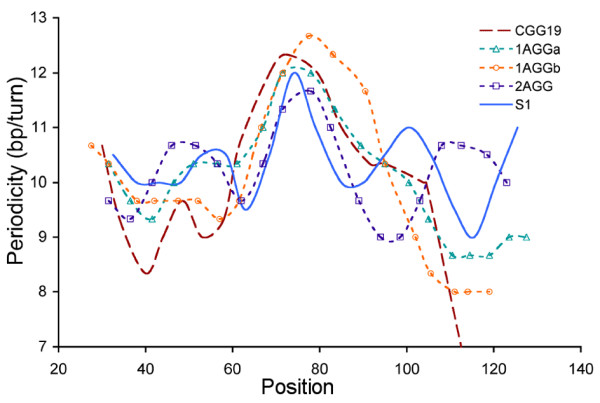
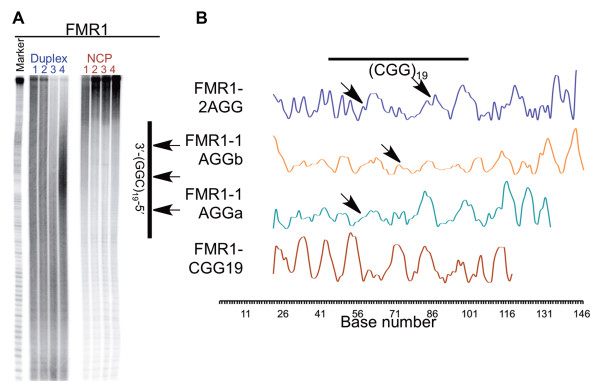
Results: To investigate how uninterrupted and interrupted CGG/CCG repeat tracts interact with the histone core, we designed experiments using the nucleosome core particle, the most basic unit of chromatin packaging. Using DNA containing 19 CGG/CCG repeats, flanked by either a nucleosome positioning sequence or the FMR1 gene sequence, we determined that the addition of a single AGG/CCT interruption modulates both the ability of the CGG/CCG repeat DNA to incorporate into a nucleosome and the rotational and translational position of the repeat DNA around the histone core when flanked by the nucleosome positioning sequence. The presence of these interruptions also alters the periodicity of the DNA in the nucleosome; interrupted repeat tracts have a greater periodicity than uninterrupted repeats.
Conclusions: This work defines the ability of AGG/CCT interruptions to modulate the behavior of the repeat tract in the packaged gene and contributes to our understanding of the role that AGG/CCT interruptions play in suppressing expansion and maintaining the correct functional organization of the FMR1 gene, highlighting a protective role played by the interruptions in genomic packaging.
Figures
Similar articles
-
Effect of CAT or AGG interruptions and CpG methylation on nucleosome assembly upon trinucleotide repeats on spinocerebellar ataxia, type 1 and fragile X syndrome.J Biol Chem. 2005 Feb 11;280(6):4498-503. doi: 10.1074/jbc.M413239200. Epub 2004 Dec 1. J Biol Chem. 2005. PMID: 15574425
-
Nucleosomal occupancy and CGG repeat expansion: a comparative analysis of triplet repeat region from mouse and human fragile X mental retardation gene 1.Chromosome Res. 2011 May;19(4):445-55. doi: 10.1007/s10577-011-9206-7. Epub 2011 Apr 16. Chromosome Res. 2011. PMID: 21499798
-
Structural Dynamics Role of AGG Interruptions in Inhibition CGG Repeat Expansion Associated with Fragile X Syndrome.ACS Chem Neurosci. 2024 Jan 17;15(2):230-235. doi: 10.1021/acschemneuro.3c00712. Epub 2023 Dec 22. ACS Chem Neurosci. 2024. PMID: 38133821
-
Contribution of DNA/RNA Structures Formed by Expanded CGG/CCG Repeats Within the FMR1 Locus in the Pathogenesis of Fragile X-Associated Disorders.Wiley Interdiscip Rev RNA. 2024 Nov-Dec;15(6):e1874. doi: 10.1002/wrna.1874. Wiley Interdiscip Rev RNA. 2024. PMID: 39523485 Review.
-
[Chromatin changes caused by CGG repeat expansion in fmr1 gene].Mol Biol (Mosk). 2015 Mar-Apr;49(2):205-11. Mol Biol (Mosk). 2015. PMID: 26065250 Review. Russian.
Cited by
-
Detection of Cryptic Fragile X Full Mutation Alleles by Southern Blot in a Female and Her Foetal DNA via Chorionic Villus Sampling, Complicated by Mosaicism for 45,X0/46,XX/47,XXX.Genes (Basel). 2021 May 24;12(6):798. doi: 10.3390/genes12060798. Genes (Basel). 2021. PMID: 34073864 Free PMC article.
-
De novo transcriptome assembly, gene annotation, and EST-SSR marker development of an important medicinal and edible crop, Amomum tsaoko (Zingiberaceae).BMC Plant Biol. 2022 Sep 29;22(1):467. doi: 10.1186/s12870-022-03827-y. BMC Plant Biol. 2022. PMID: 36171538 Free PMC article.
-
The association of CGG repeat length and AGG interruption patterns on FMR1 alleles with female infertility.Front Endocrinol (Lausanne). 2025 Jun 17;16:1609471. doi: 10.3389/fendo.2025.1609471. eCollection 2025. Front Endocrinol (Lausanne). 2025. PMID: 40600017 Free PMC article.
-
Expansions and contractions of the FMR1 CGG repeat in 5,508 transmissions of normal, intermediate, and premutation alleles.Am J Med Genet A. 2019 Jul;179(7):1148-1156. doi: 10.1002/ajmg.a.61165. Epub 2019 May 2. Am J Med Genet A. 2019. PMID: 31050164 Free PMC article.
-
Defining the role of the CGGBP1 protein in FMR1 gene expression.Eur J Hum Genet. 2016 May;24(5):697-703. doi: 10.1038/ejhg.2015.182. Epub 2015 Aug 26. Eur J Hum Genet. 2016. PMID: 26306647 Free PMC article.
References
-
- Verkerk AJ, Pieretti M, Sutcliffe JS, Fu YH, Kuhl DP, Pizzuti A, Reiner O, Richards S, Victoria MF, Zhang FP. Identification of a gene (FMR-1) containing a CGG repeat coincident with a breakpoint cluster region exhibiting length variation in fragile X syndrome. Cell. 1991;65:905–914. - PubMed
-
- Fu YH, Kuhl DP, Pizzuti A, Pieretti M, Sutcliffe JS, Richards S, Verkerk AJ, Holden JJ, Fenwick RG, Warren ST. Variation of the CGG repeat at the fragile X site results in genetic instability: resolution of the Sherman paradox. Cell. 1991;67:1047–1058. - PubMed
-
- Oberle I, Rousseau F, Heitz D, Kretz C, Devys D, Hanauer A, Boue J, Bertheas M, Mandel J. Instability of a 550-base pair DNA segment and abnormal methylation in fragile X syndrome. Science. 1991;252:1097–1102. - PubMed
-
- Heitz D, Rousseau F, Devys D, Saccone S, Abderrahim H, Le Paslier D, Cohen D, Vincent A, Toniolo D, Valle Della G. Isolation of sequences that span the fragile X and identification of a fragile X-related CpG island. Science. 1991;251:1236–1239. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
