

Theoretical foundations of the sound analog membrane potential that underlies coincidence detection in the barn owl
- PMID: 24265616
- PMCID: PMC3821005
- DOI: 10.3389/fncom.2013.00151
Theoretical foundations of the sound analog membrane potential that underlies coincidence detection in the barn owl
Abstract
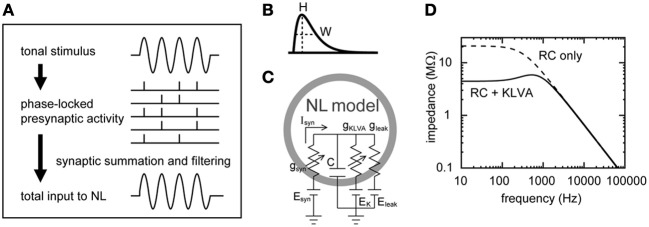
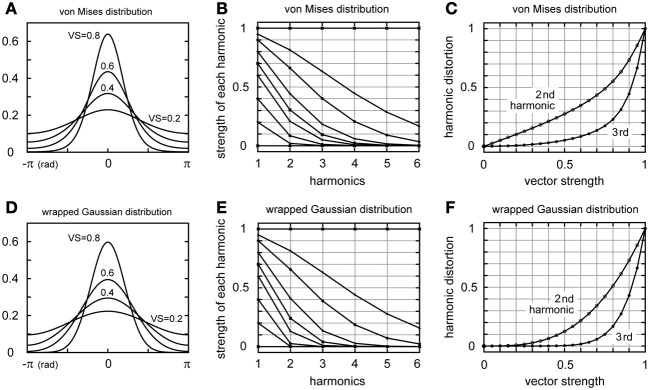
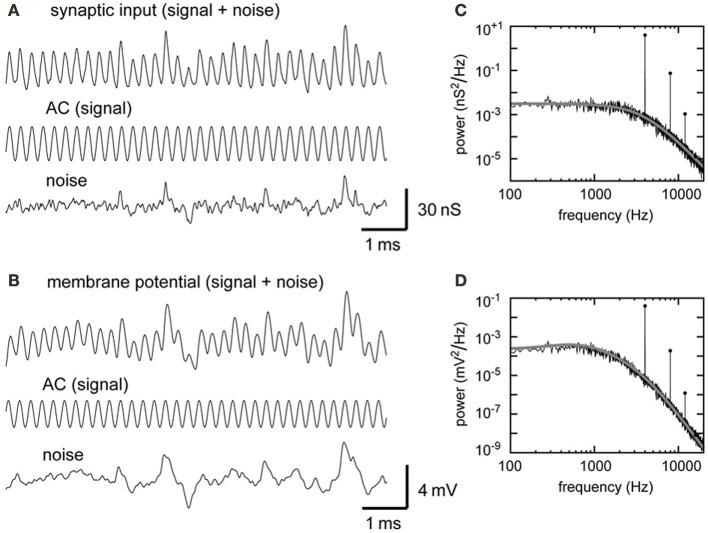
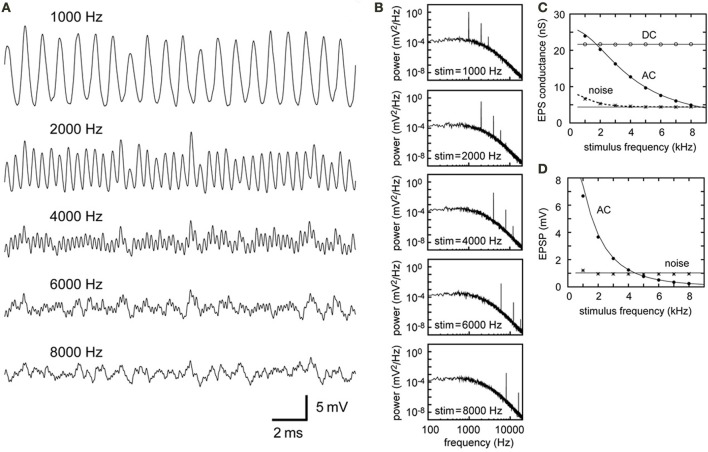
A wide variety of neurons encode temporal information via phase-locked spikes. In the avian auditory brainstem, neurons in the cochlear nucleus magnocellularis (NM) send phase-locked synaptic inputs to coincidence detector neurons in the nucleus laminaris (NL) that mediate sound localization. Previous modeling studies suggested that converging phase-locked synaptic inputs may give rise to a periodic oscillation in the membrane potential of their target neuron. Recent physiological recordings in vivo revealed that owl NL neurons changed their spike rates almost linearly with the amplitude of this oscillatory potential. The oscillatory potential was termed the sound analog potential, because of its resemblance to the waveform of the stimulus tone. The amplitude of the sound analog potential recorded in NL varied systematically with the interaural time difference (ITD), which is one of the most important cues for sound localization. In order to investigate the mechanisms underlying ITD computation in the NM-NL circuit, we provide detailed theoretical descriptions of how phase-locked inputs form oscillating membrane potentials. We derive analytical expressions that relate presynaptic, synaptic, and postsynaptic factors to the signal and noise components of the oscillation in both the synaptic conductance and the membrane potential. Numerical simulations demonstrate the validity of the theoretical formulations for the entire frequency ranges tested (1-8 kHz) and potential effects of higher harmonics on NL neurons with low best frequencies (<2 kHz).
Keywords: auditory brainstem; oscillation; owl; periodic signals; phase-locking; sound localization.
Figures
References
-
- Abramowitz M., Stegun I. A. (eds.). (1972). Handbook of Mathematical Functions with Formulas, Graphs, and Mathematical Tables (tenth printing). New York, NY: Dover
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources