

PrimPol bypasses UV photoproducts during eukaryotic chromosomal DNA replication
- PMID: 24267451
- PMCID: PMC4228047
- DOI: 10.1016/j.molcel.2013.10.035
PrimPol bypasses UV photoproducts during eukaryotic chromosomal DNA replication
Abstract
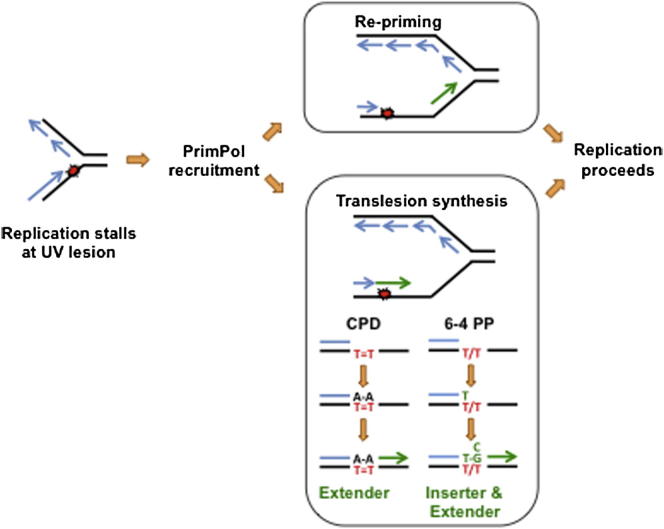
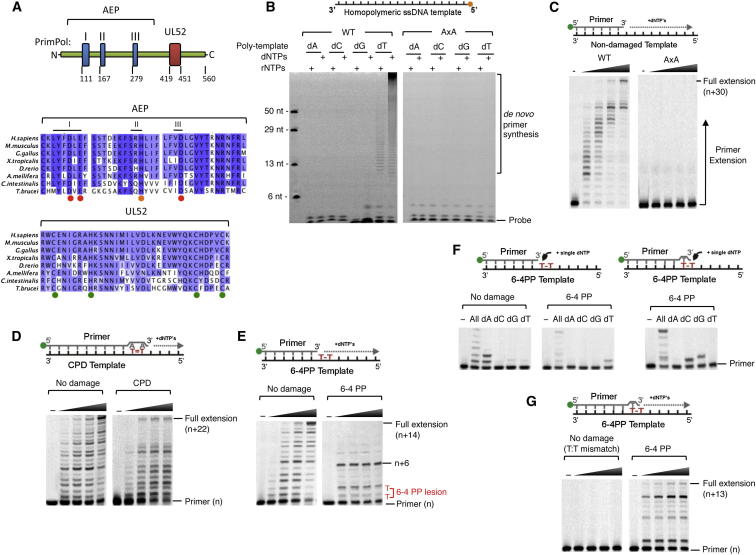
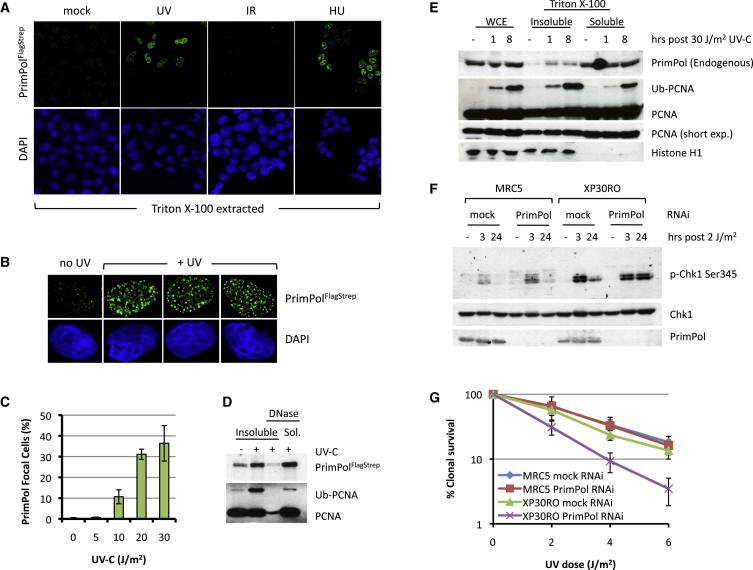
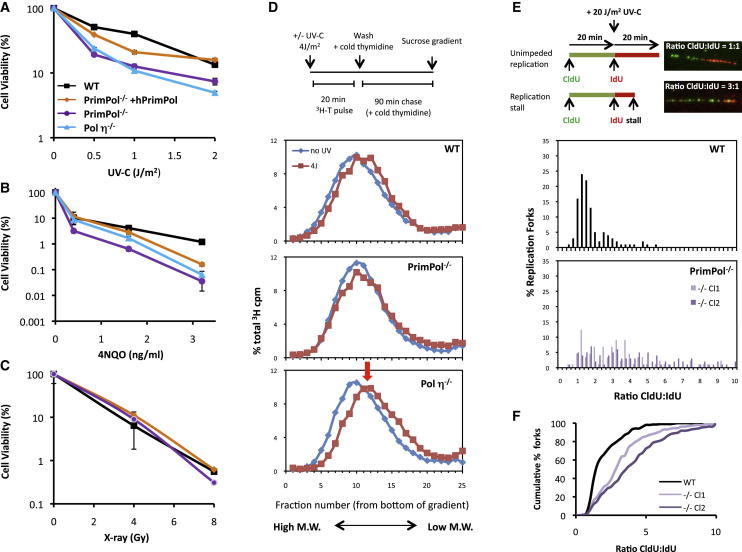
DNA damage can stall the DNA replication machinery, leading to genomic instability. Thus, numerous mechanisms exist to complete genome duplication in the absence of a pristine DNA template, but identification of the enzymes involved remains incomplete. Here, we establish that Primase-Polymerase (PrimPol; CCDC111), an archaeal-eukaryotic primase (AEP) in eukaryotic cells, is involved in chromosomal DNA replication. PrimPol is required for replication fork progression on ultraviolet (UV) light-damaged DNA templates, possibly mediated by its ability to catalyze translesion synthesis (TLS) of these lesions. This PrimPol UV lesion bypass pathway is not epistatic with the Pol η-dependent pathway and, as a consequence, protects xeroderma pigmentosum variant (XP-V) patient cells from UV-induced cytotoxicity. In addition, we establish that PrimPol is also required for efficient replication fork progression during an unperturbed S phase. These and other findings indicate that PrimPol is an important player in replication fork progression in eukaryotic cells.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
References
-
- Aguilera A., Gómez-González B. Genome instability: a mechanistic view of its causes and consequences. Nat. Rev. Genet. 2008;9:204–217. - PubMed
-
- Arlett C.F., Harcourt S.A., Broughton B.C. The influence of caffeine on cell survival in excision-proficient and excision-deficient xeroderma pigmentosum and normal human cell strains following ultraviolet-light irradiation. Mutat. Res. 1975;33:341–346. - PubMed
-
- Bocquier A.A., Liu L., Cann I.K., Komori K., Kohda D., Ishino Y. Archaeal primase: bridging the gap between RNA and DNA polymerases. Curr. Biol. 2001;11:452–456. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
