

Phosphorylation-regulated binding of RNA polymerase II to fibrous polymers of low-complexity domains
- PMID: 24267890
- PMCID: PMC4010232
- DOI: 10.1016/j.cell.2013.10.033
Phosphorylation-regulated binding of RNA polymerase II to fibrous polymers of low-complexity domains
Erratum in
- Cell. 2014 Jan 16;156(1-2):374
Abstract
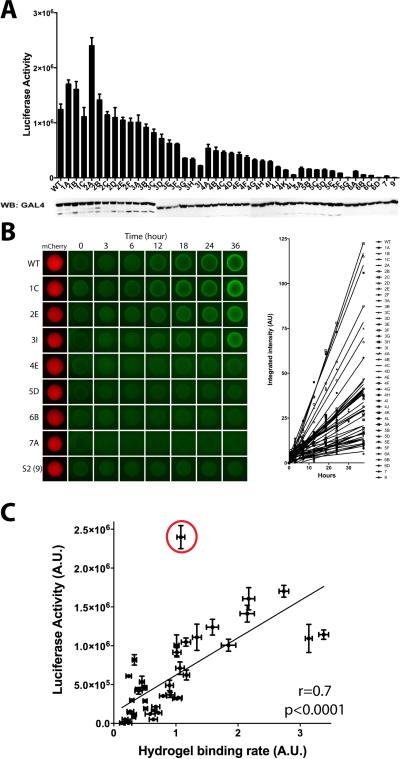
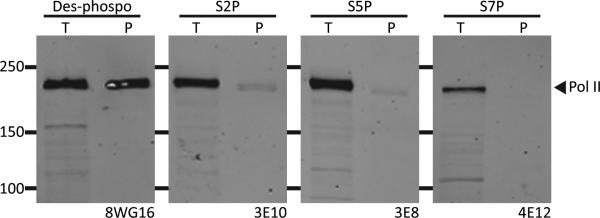
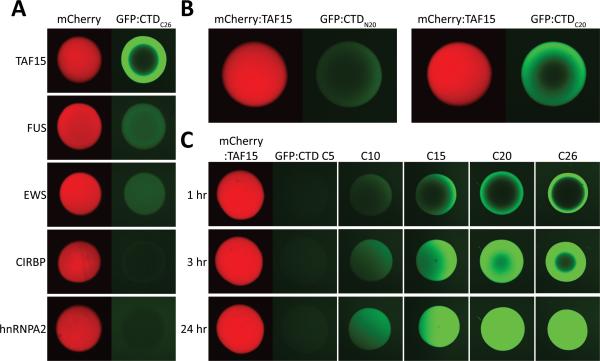
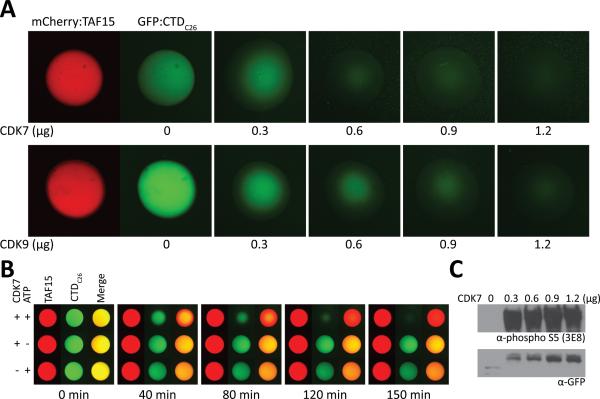
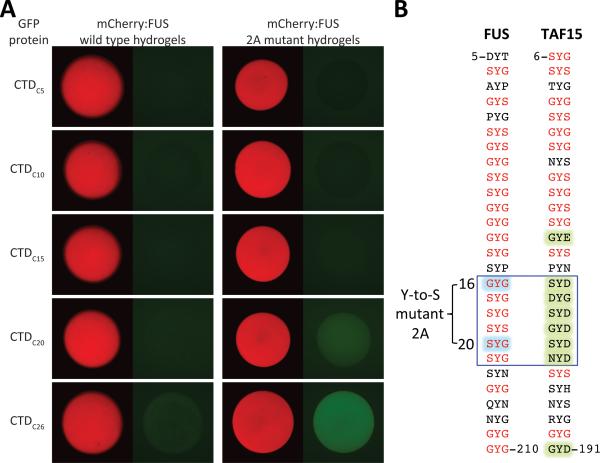
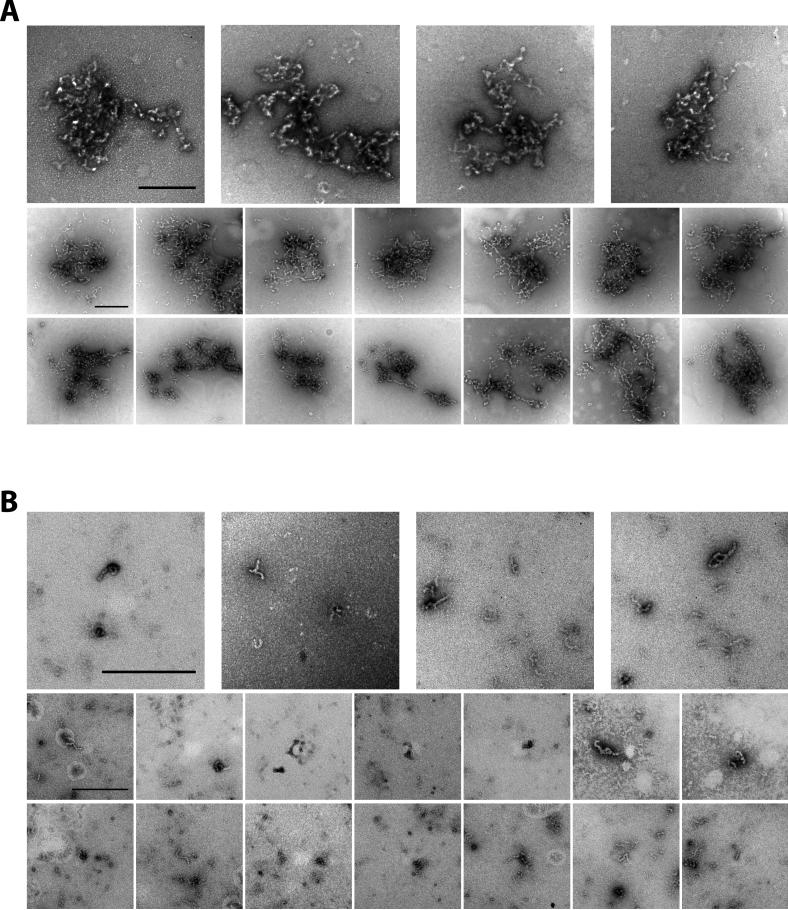
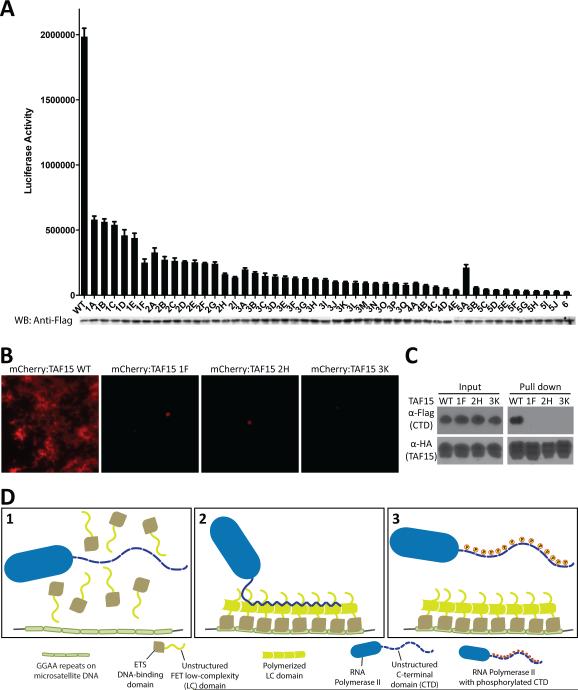
The low-complexity (LC) domains of the products of the fused in sarcoma (FUS), Ewings sarcoma (EWS), and TAF15 genes are translocated onto a variety of different DNA-binding domains and thereby assist in driving the formation of cancerous cells. In the context of the translocated fusion proteins, these LC sequences function as transcriptional activation domains. Here, we show that polymeric fibers formed from these LC domains directly bind the C-terminal domain (CTD) of RNA polymerase II in a manner reversible by phosphorylation of the iterated, heptad repeats of the CTD. Mutational analysis indicates that the degree of binding between the CTD and the LC domain polymers correlates with the strength of transcriptional activation. These studies offer a simple means of conceptualizing how RNA polymerase II is recruited to active genes in its unphosphorylated state and released for elongation following phosphorylation of the CTD.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
