

Mapping the primate lateral geniculate nucleus: a review of experiments and methods
- PMID: 24270042
- PMCID: PMC5446894
- DOI: 10.1016/j.jphysparis.2013.10.001
Mapping the primate lateral geniculate nucleus: a review of experiments and methods
Abstract
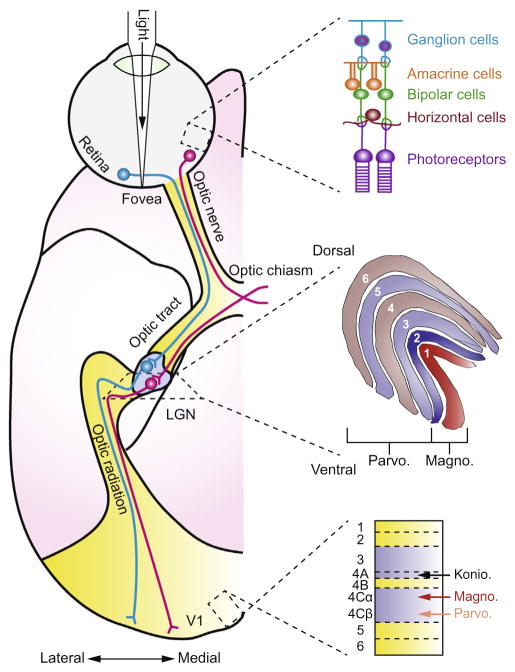
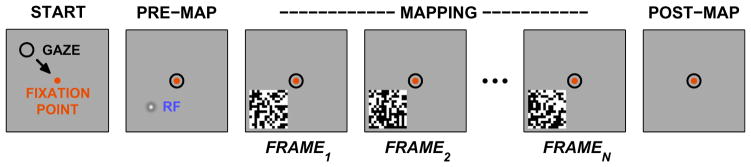
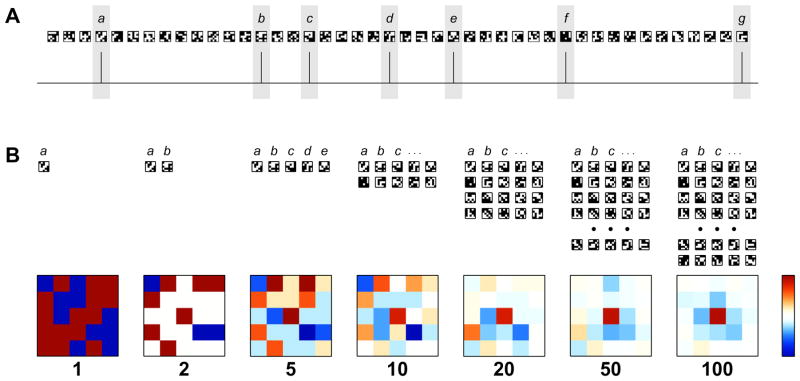
Mapping neuronal responses in the lateral geniculate nucleus (LGN) is key to understanding how visual information is processed in the brain. This paper focuses on our current knowledge of the dynamics the receptive field (RF) as broken down into the classical receptive field (CRF) and the extra-classical receptive field (ECRF) in primate LGN. CRFs in the LGN are known to be similar to those in the retinal ganglion cell layer in terms of both spatial and temporal characteristics, leading to the standard interpretation of the LGN as a relay center from retina to primary visual cortex. ECRFs have generally been found to be large and inhibitory, with some differences in magnitude between the magno-, parvo-, and koniocellular pathways. The specific contributions of the retina, thalamus, and visual cortex to LGN ECRF properties are presently unknown. Some reports suggest a retinal origin for extra-classical suppression based on latency arguments and other reports have suggested a thalamic origin for extra-classical suppression. This issue is complicated by the use of anesthetized animals, where cortical activity is likely to be altered. Thus further study of LGN ECRFs is warranted to reconcile these discrepancies. Producing descriptions of RF properties of LGN neurons could be enhanced by employing preferred naturalistic stimuli. Although there has been significant work in cats with natural scene stimuli and noise that statistically imitates natural scenes, we highlight a need for similar data from primates. Obtaining these data may be aided by recent advancements in experimental and analytical techniques that permit the efficient study of nonlinear RF characteristics in addition to traditional linear factors. In light of the reviewed topics, we conclude by suggesting experiments to more clearly elucidate the spatial and temporal structure of ECRFs of primate LGN neurons.
Keywords: CRF; EC; ECI; ECRF; Early visual system; Extra-classical receptive field; K; LGN; M; MID; Maximally Informative Dimensions; P; Parvocellular; RF; RGC; Reverse correlation; STA; STC; Thalamus; V1; classical receptive field; extra-classical; extra-classical inhibition; extra-classical receptive field; koniocellular; lateral geniculate nucleus; magnocellular; primary visual cortex; receptive field; retinal ganglion cell; spike-triggered average; spike-triggered covariance.
Copyright © 2013 The Authors. Published by Elsevier Ltd.. All rights reserved.
Figures

References
-
- Angelucci A, Bressloff PC. Contribution of feedforward, lateral and feedback connections to the classical receptive field center and extra-classical receptive field surround of primate V1 neurons. Prog Brain Res. 2006:93–120. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
