

ADCK4 mutations promote steroid-resistant nephrotic syndrome through CoQ10 biosynthesis disruption
- PMID: 24270420
- PMCID: PMC3859425
- DOI: 10.1172/JCI69000
ADCK4 mutations promote steroid-resistant nephrotic syndrome through CoQ10 biosynthesis disruption
Abstract
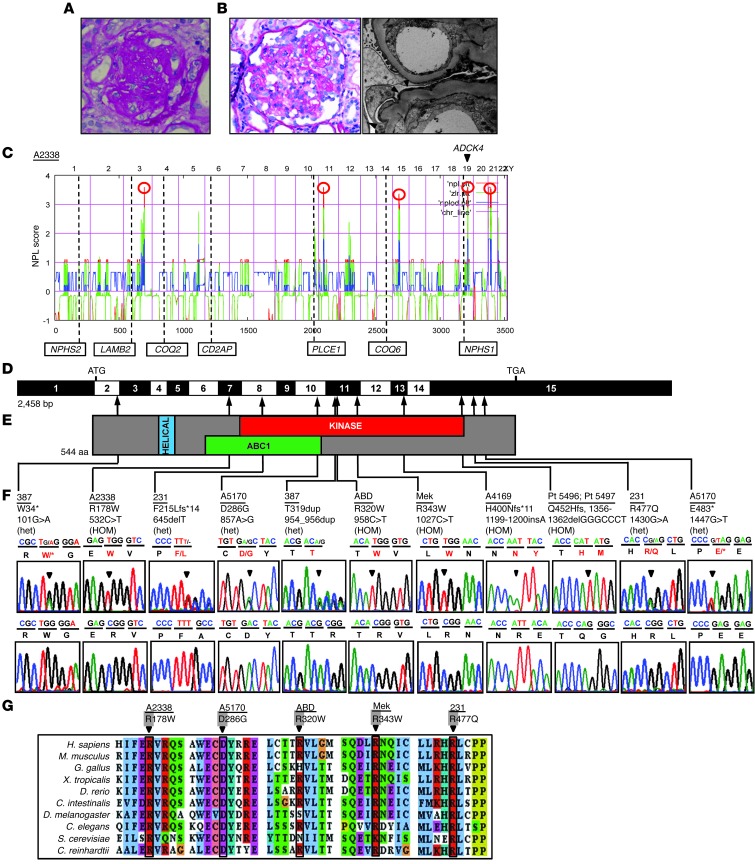
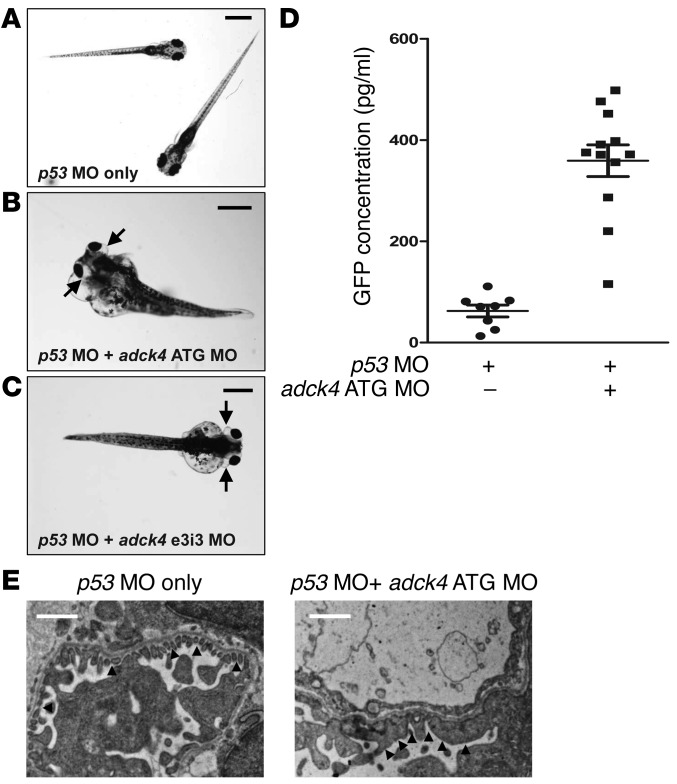
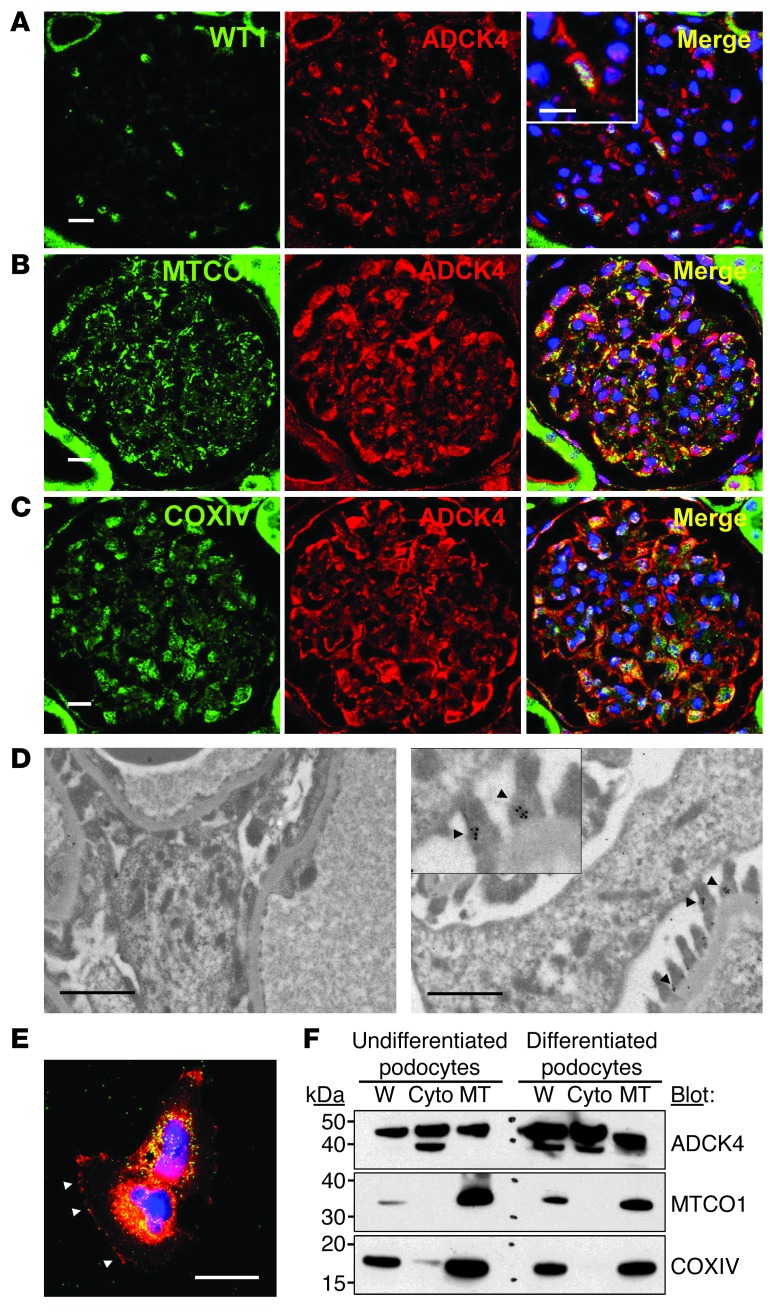
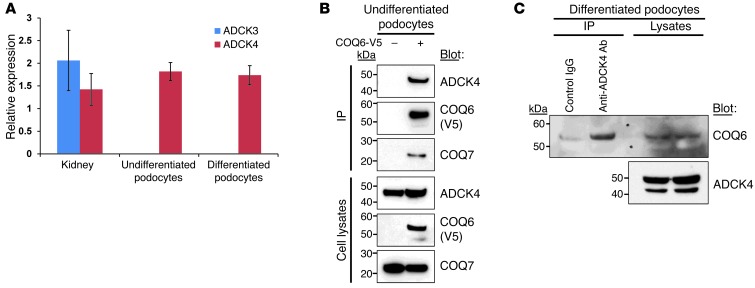
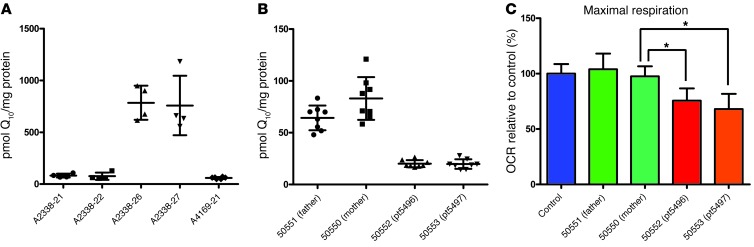
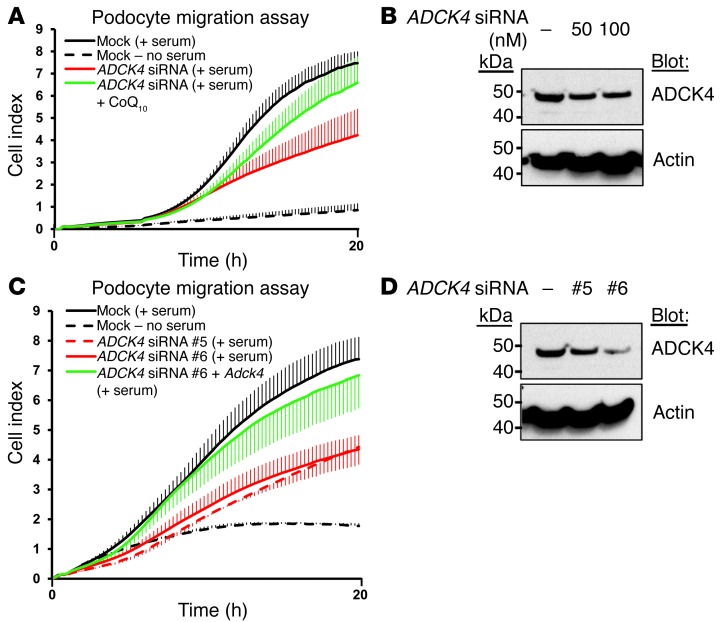
Identification of single-gene causes of steroid-resistant nephrotic syndrome (SRNS) has furthered the understanding of the pathogenesis of this disease. Here, using a combination of homozygosity mapping and whole human exome resequencing, we identified mutations in the aarF domain containing kinase 4 (ADCK4) gene in 15 individuals with SRNS from 8 unrelated families. ADCK4 was highly similar to ADCK3, which has been shown to participate in coenzyme Q10 (CoQ10) biosynthesis. Mutations in ADCK4 resulted in reduced CoQ10 levels and reduced mitochondrial respiratory enzyme activity in cells isolated from individuals with SRNS and transformed lymphoblasts. Knockdown of adck4 in zebrafish and Drosophila recapitulated nephrotic syndrome-associated phenotypes. Furthermore, ADCK4 was expressed in glomerular podocytes and partially localized to podocyte mitochondria and foot processes in rat kidneys and cultured human podocytes. In human podocytes, ADCK4 interacted with members of the CoQ10 biosynthesis pathway, including COQ6, which has been linked with SRNS and COQ7. Knockdown of ADCK4 in podocytes resulted in decreased migration, which was reversed by CoQ10 addition. Interestingly, a patient with SRNS with a homozygous ADCK4 frameshift mutation had partial remission following CoQ10 treatment. These data indicate that individuals with SRNS with mutations in ADCK4 or other genes that participate in CoQ10 biosynthesis may be treatable with CoQ10.
Figures
Comment in
-
ADCK4 "reenergizes" nephrotic syndrome.J Clin Invest. 2013 Dec;123(12):4996-9. doi: 10.1172/JCI73168. Epub 2013 Nov 25. J Clin Invest. 2013. PMID: 24270414 Free PMC article.
References
-
- Kim MS, Stablein D, Harmon WE. Renal transplantation in children with congenital nephrotic syndrome: a report of the North American Pediatric Renal Transplant Cooperative Study (NAPRTCS). Pediatr Transplant. 1998;2(4):305–308. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 DK054931/DK/NIDDK NIH HHS/United States
- R01 EY007042/EY/NEI NIH HHS/United States
- R01 DK076683/DK/NIDDK NIH HHS/United States
- R01HL090801/HL/NHLBI NIH HHS/United States
- GM 007185/GM/NIGMS NIH HHS/United States
- T32 GM007185/GM/NIGMS NIH HHS/United States
- DK090917/DK/NIDDK NIH HHS/United States
- DK086542/DK/NIDDK NIH HHS/United States
- R01 EY013408/EY/NEI NIH HHS/United States
- G0800571/MRC_/Medical Research Council/United Kingdom
- R01 DK046073/DK/NIDDK NIH HHS/United States
- DK091405/DK/NIDDK NIH HHS/United States
- DK076683/DK/NIDDK NIH HHS/United States
- RC1 DK086542/DK/NIDDK NIH HHS/United States
- DK46073/DK/NIDDK NIH HHS/United States
- DK081943/DK/NIDDK NIH HHS/United States
- R56 DK046073/DK/NIDDK NIH HHS/United States
- P30 DK079310/DK/NIDDK NIH HHS/United States
- U54HG006504/HG/NHGRI NIH HHS/United States
- U54 HG006504/HG/NHGRI NIH HHS/United States
- R00 DK091405/DK/NIDDK NIH HHS/United States
- S10RR024605/RR/NCRR NIH HHS/United States
- R01 HL090801/HL/NHLBI NIH HHS/United States
- S10 RR024605/RR/NCRR NIH HHS/United States
- P30 DK081943/DK/NIDDK NIH HHS/United States
- RC4 DK090917/DK/NIDDK NIH HHS/United States
- EY07042/EY/NEI NIH HHS/United States
- K99 DK091405/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
