

The human cap-binding complex is functionally connected to the nuclear RNA exosome
- PMID: 24270879
- PMCID: PMC3923317
- DOI: 10.1038/nsmb.2703
The human cap-binding complex is functionally connected to the nuclear RNA exosome
Abstract
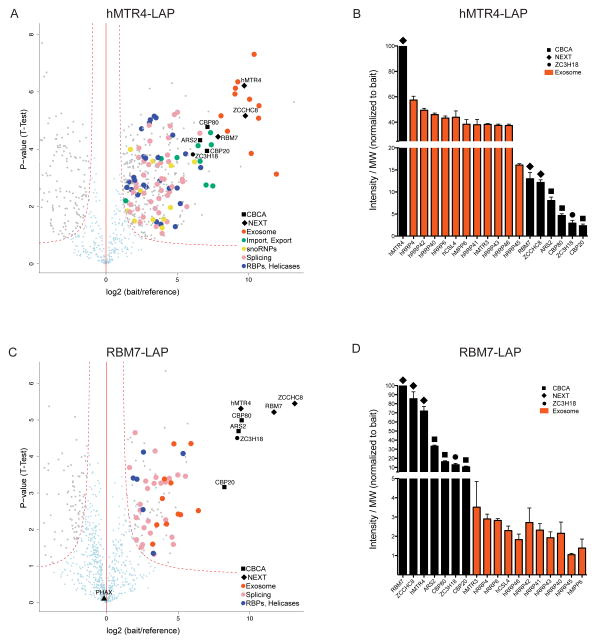
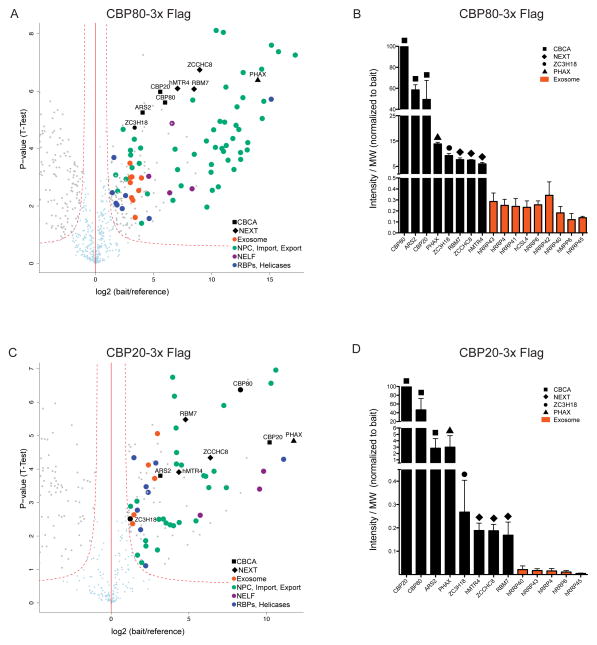
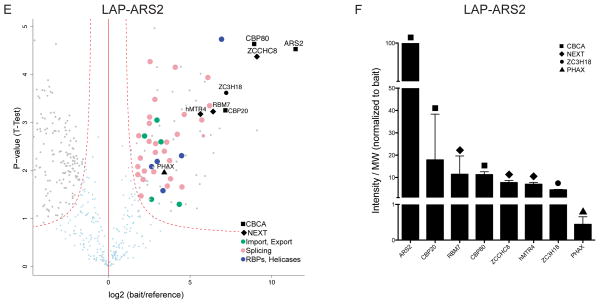
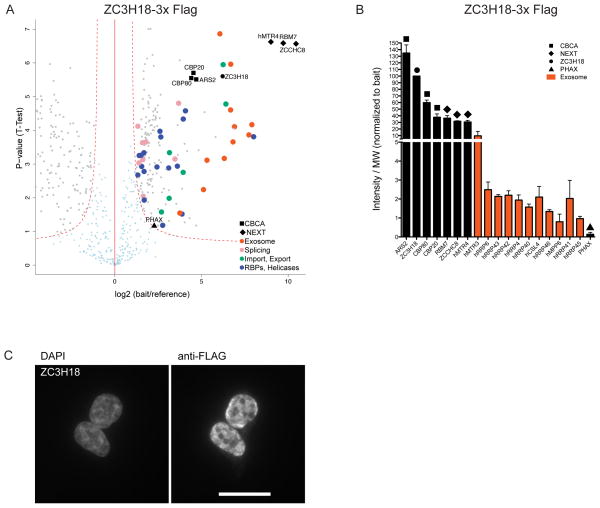
Nuclear processing and quality control of eukaryotic RNA is mediated by the RNA exosome, which is regulated by accessory factors. However, the mechanism of exosome recruitment to its ribonucleoprotein (RNP) targets remains poorly understood. Here we report a physical link between the human exosome and the cap-binding complex (CBC). The CBC associates with the ARS2 protein to form CBC-ARS2 (CBCA) and then further connects, together with the ZC3H18 protein, to the nuclear exosome targeting (NEXT) complex, thus forming CBC-NEXT (CBCN). RNA immunoprecipitation using CBCN factors as well as the analysis of combinatorial depletion of CBCN and exosome components underscore the functional relevance of CBC-exosome bridging at the level of target RNA. Specifically, CBCA suppresses read-through products of several RNA families by promoting their transcriptional termination. We suggest that the RNP 5' cap links transcription termination to exosomal RNA degradation through CBCN.
Figures
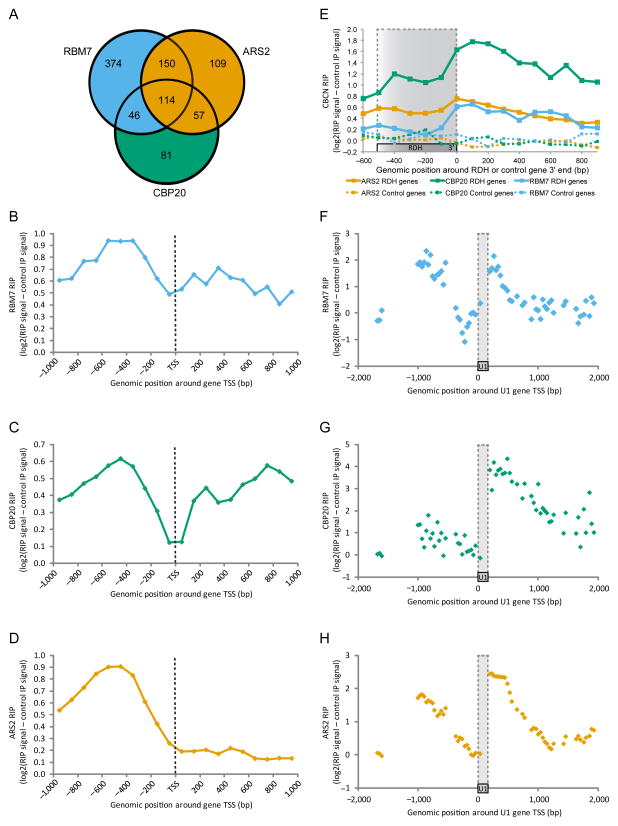
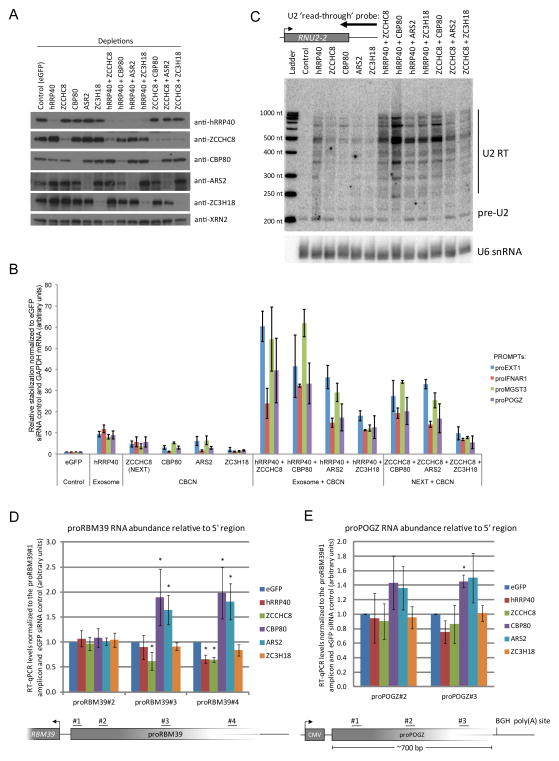
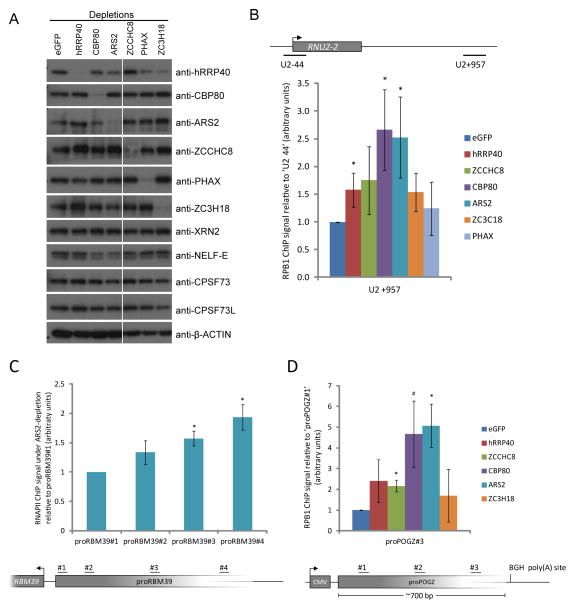
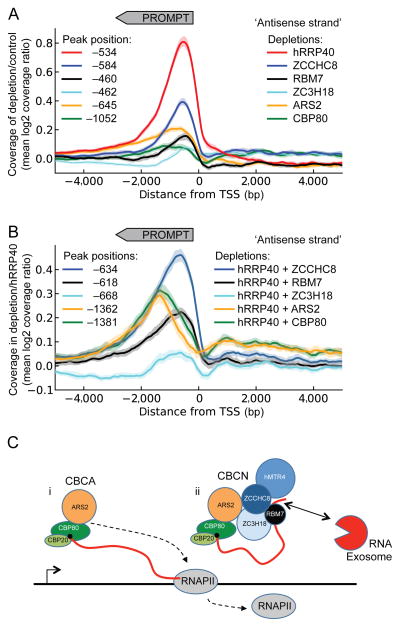
Comment in
-
Good cap/bad cap: how the cap-binding complex determines RNA fate.Nat Struct Mol Biol. 2014 Jan;21(1):9-12. doi: 10.1038/nsmb.2751. Nat Struct Mol Biol. 2014. PMID: 24389545 No abstract available.
References
-
- Chlebowski A, Lubas M, Jensen TH, Dziembowski A. RNA decay machines: The exosome. Biochim Biophys Acta. 2013;1829:552–60. - PubMed
-
- Houseley J, Tollervey D. The many pathways of RNA degradation. Cell. 2009;136:763–76. - PubMed
-
- Lykke-Andersen S, Brodersen DE, Jensen TH. Origins and activities of the eukaryotic exosome. J Cell Sci. 2009;122:1487–94. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- SRA/SRP031620
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
