

Organizing principles of mammalian nonsense-mediated mRNA decay
- PMID: 24274751
- PMCID: PMC4148824
- DOI: 10.1146/annurev-genet-111212-133424
Organizing principles of mammalian nonsense-mediated mRNA decay
Abstract
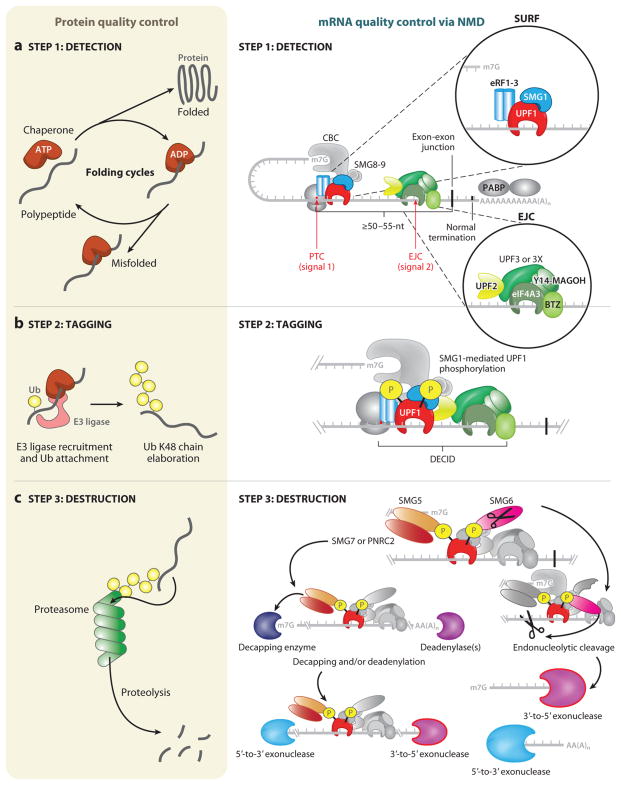
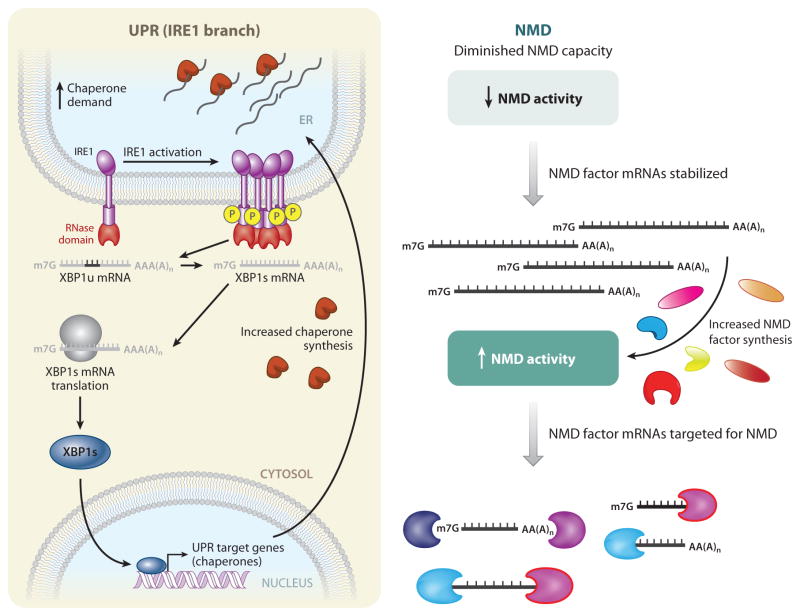
Cells use messenger RNAs (mRNAs) to ensure the accurate dissemination of genetic information encoded by DNA. Given that mRNAs largely direct the synthesis of a critical effector of cellular phenotype, i.e., proteins, tight regulation of both the quality and quantity of mRNA is a prerequisite for effective cellular homeostasis. Here, we review nonsense-mediated mRNA decay (NMD), which is the best-characterized posttranscriptional quality control mechanism that cells have evolved in their cytoplasm to ensure transcriptome fidelity. We use protein quality control as a conceptual framework to organize what is known about NMD, highlighting overarching similarities between these two polymer quality control pathways, where the protein quality control and NMD pathways intersect, and how protein quality control can suggest new avenues for research into mRNA quality control.
Figures
References
-
- Amrani N, Ganesan R, Kervestin S, Mangus DA, Ghosh S, Jacobson A. A faux 3′-UTR promotes aberrant termination and triggers nonsense-mediated mRNA decay. Nature. 2004;432:112–18. - PubMed
-
- Ballut L, Marchadier B, Baguet A, Tomasetto C, Séraphin B, Le Hir H. The exon junction core complex is locked onto RNA by inhibition of eIF4AIII ATPase activity. Nat Struct Mol Biol. 2005;12:861–69. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
