

The monogenean which lost its clamps
- PMID: 24278118
- PMCID: PMC3838368
- DOI: 10.1371/journal.pone.0079155
The monogenean which lost its clamps
Abstract
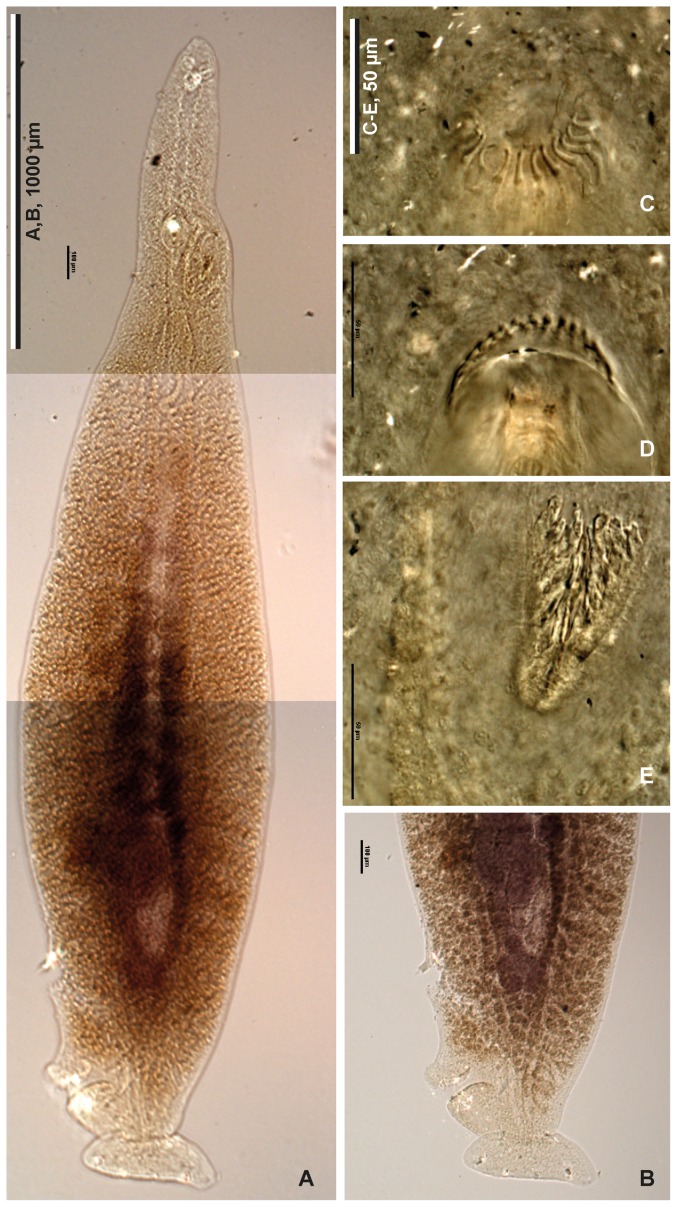
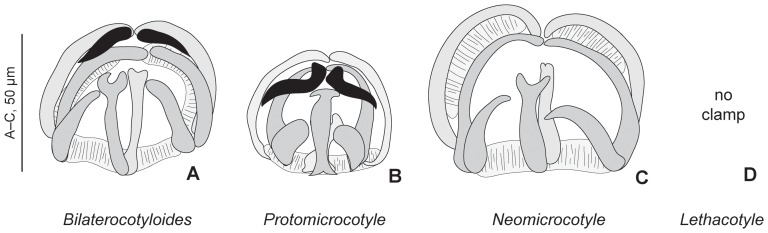
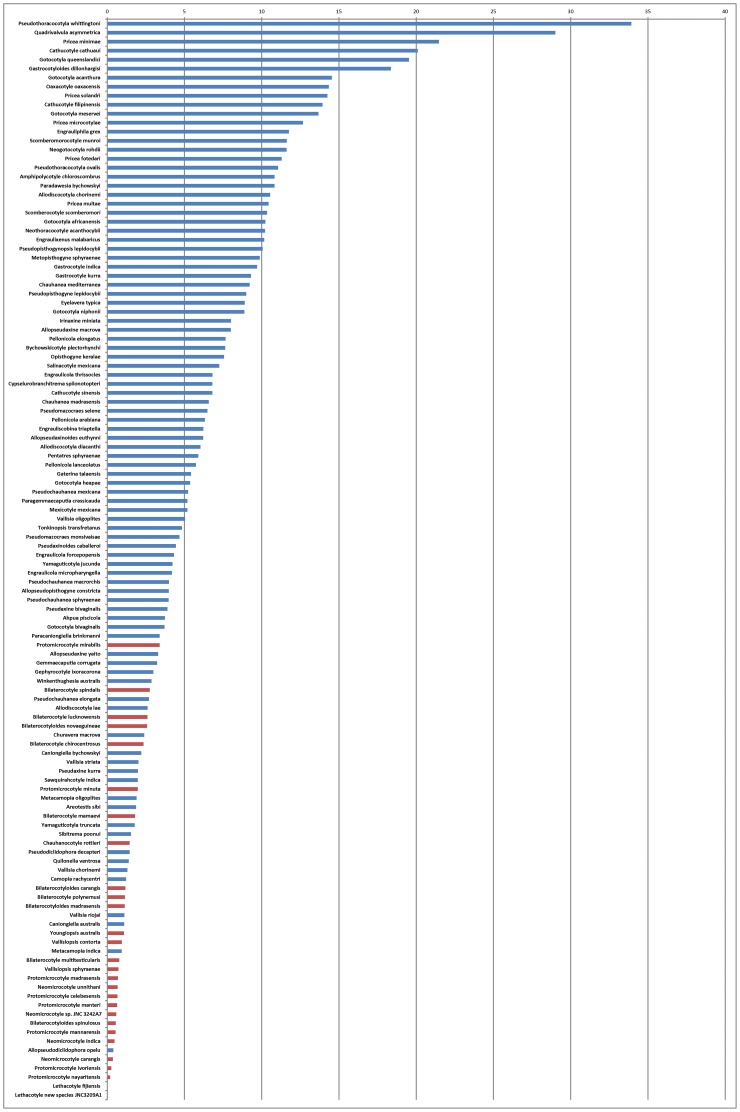
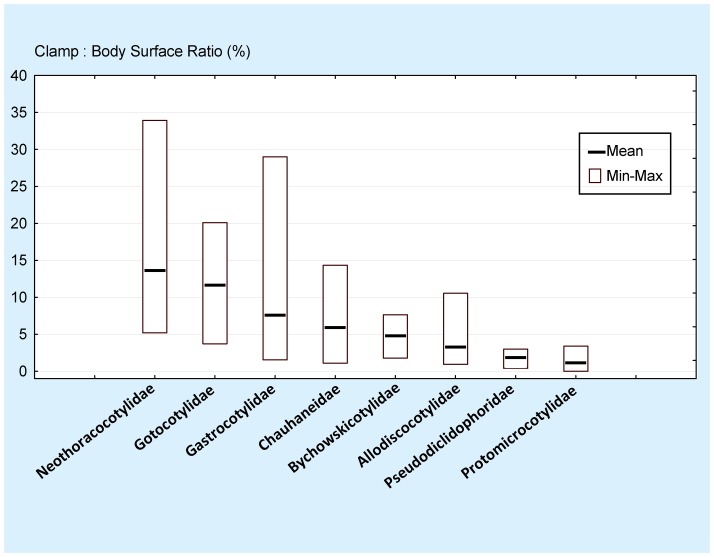
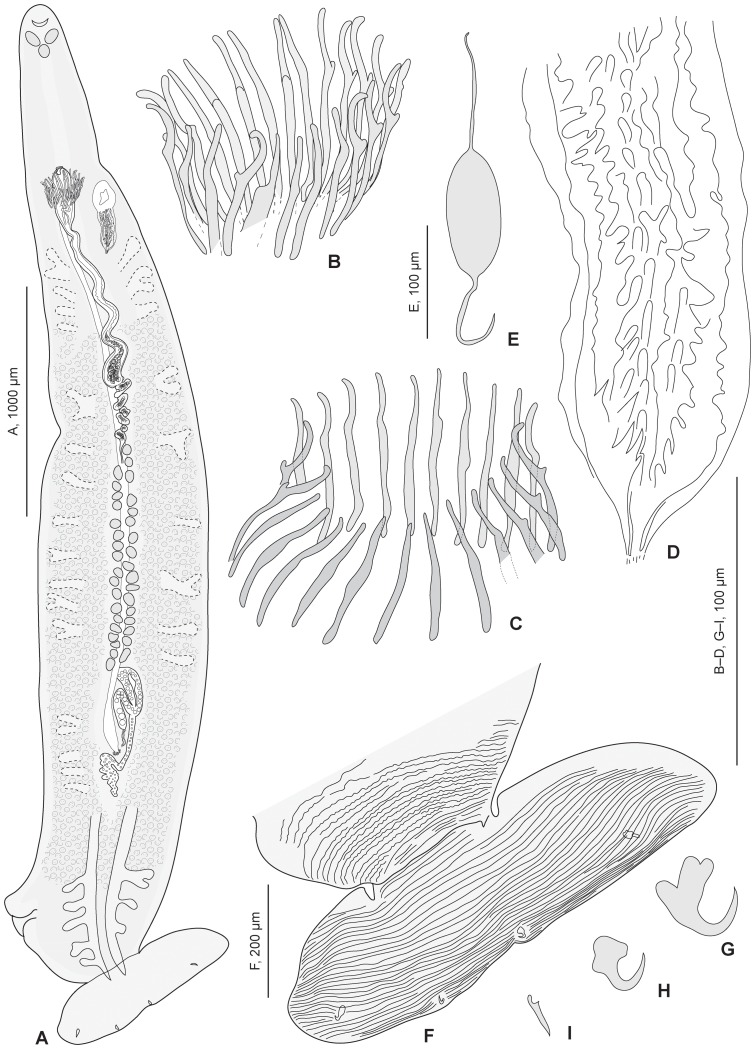
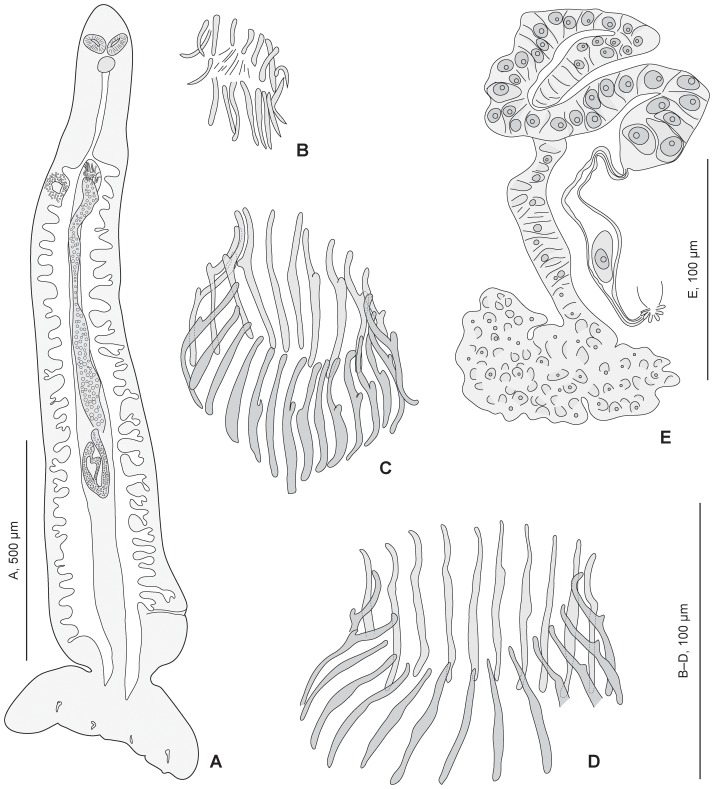
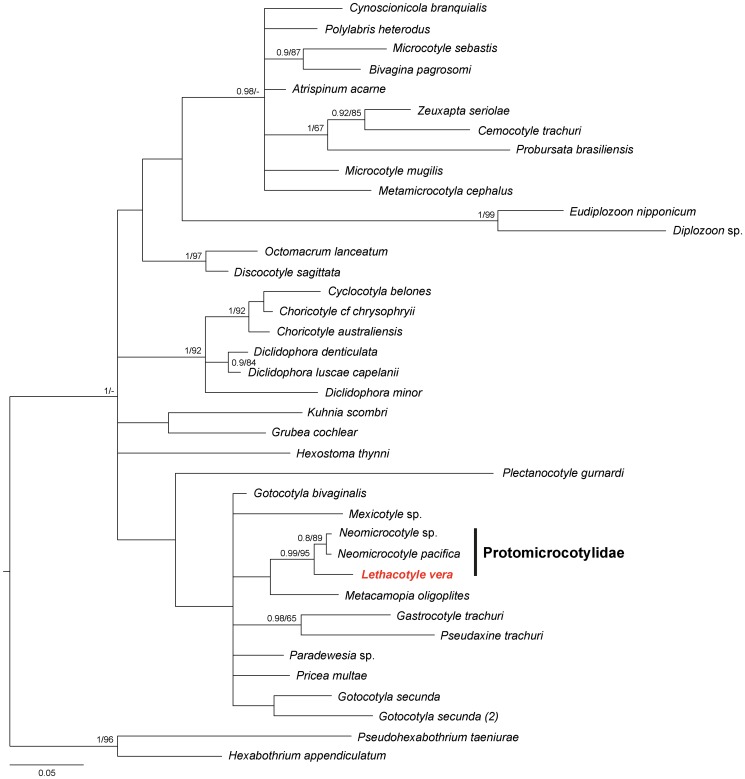
Ectoparasites face a daily challenge: to remain attached to their hosts. Polyopisthocotylean monogeneans usually attach to the surface of fish gills using highly specialized structures, the sclerotized clamps. In the original description of the protomicrocotylid species Lethacotyle fijiensis, described 60 years ago, the clamps were considered to be absent but few specimens were available and this observation was later questioned. In addition, genera within the family Protomicrocotylidae have either clamps of the "gastrocotylid" or the "microcotylid" types; this puzzled systematists because these clamp types are characteristic of distinct, major groups. Discovery of another, new, species of the genus Lethacotyle, has allowed us to explore the nature of the attachment structures in protomicrocotylids. Lethacotyle vera n. sp. is described from the gills of the carangid Caranx papuensis off New Caledonia. It is distinguished from Lethacotyle fijiensis, the only other species of the genus, by the length of the male copulatory spines. Sequences of 28S rDNA were used to build a tree, in which Lethacotyle vera grouped with other protomicrocotylids. The identity of the host fish was confirmed with COI barcodes. We observed that protomicrocotylids have specialized structures associated with their attachment organ, such as lateral flaps and transverse striations, which are not known in other monogeneans. We thus hypothesized that the clamps in protomicrocotylids were sequentially lost during evolution, coinciding with the development of other attachment structures. To test the hypothesis, we calculated the surfaces of clamps and body in 120 species of gastrocotylinean monogeneans, based on published descriptions. The ratio of clamp surface: body surface was the lowest in protomicrocotylids. We conclude that clamps in protomicrocotylids are vestigial organs, and that occurrence of "gastrocotylid" and simpler "microcotylid" clamps within the same family are steps in an evolutionary sequence, leading to the absence of these attributes in species of Lethacotyle.
Conflict of interest statement
Figures


References
-
- Justine J-L (1998) Non-monophyly of the monogeneans? International Journal for Parasitology 28: 1653–1657. - PubMed
-
- Perkins EM, Donnellan SC, Bertozzi T, Whittington ID (2010) Closing the mitochondrial circle on paraphyly of the Monogenea (Platyhelminthes) infers evolution in the diet of parasitic flatworms. International Journal for Parasitology 40: 1237–1245. - PubMed
-
- Mollaret I, Jamieson BGM, Adlard RD, Hugall A, Lecointre G, et al. (1997) Phylogenetic analysis of the Monogenea and their relationships with Digenea and Eucestoda inferred from 28S rDNA sequences. Molecular and Biochemical Parasitology 90: 433–438. - PubMed
-
- Mollaret I, Jamieson BGM, Justine J-L (2000) Phylogeny of the Monopisthocotylea and Polyopisthocotylea (Platyhelminthes) inferred from 28S rDNA sequences. International Journal for Parasitology 30: 171–185. - PubMed
-
- Littlewood DTJ, Rohde K, Clough KA (1999) The interrelationships of all major groups of Platyhelminthes: phylogenetic evidence from morphology and molecules. Biological Journal of the Linnean Society 66: 75–114.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
