

Changes in the inflammatory response to injury and its resolution during the loss of regenerative capacity in developing Xenopus limbs
- PMID: 24278286
- PMCID: PMC3835323
- DOI: 10.1371/journal.pone.0080477
Changes in the inflammatory response to injury and its resolution during the loss of regenerative capacity in developing Xenopus limbs
Abstract
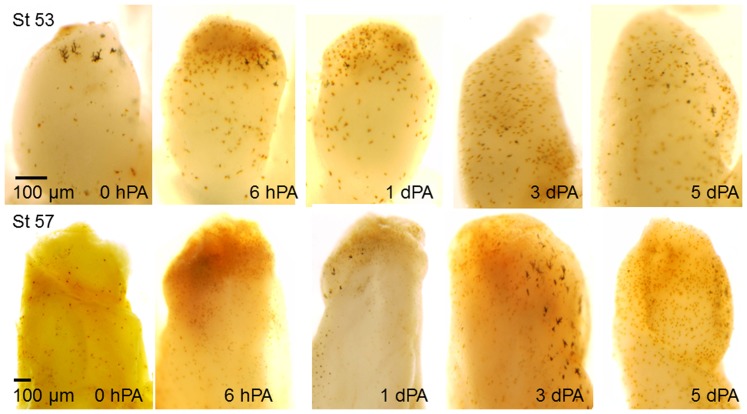
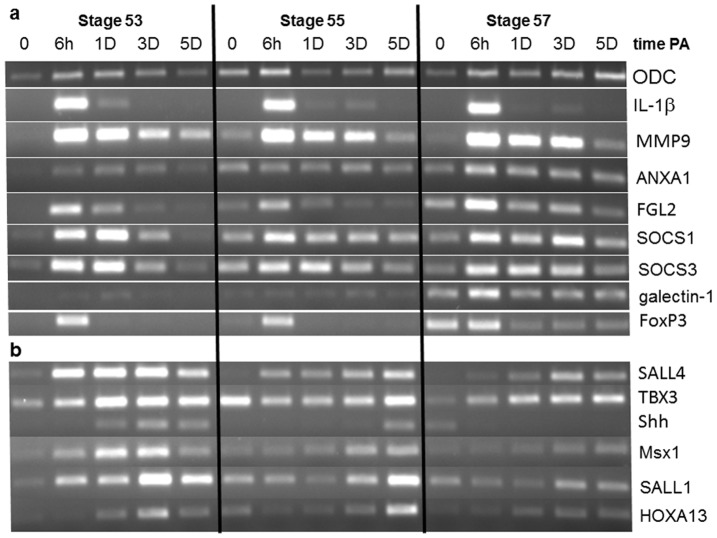
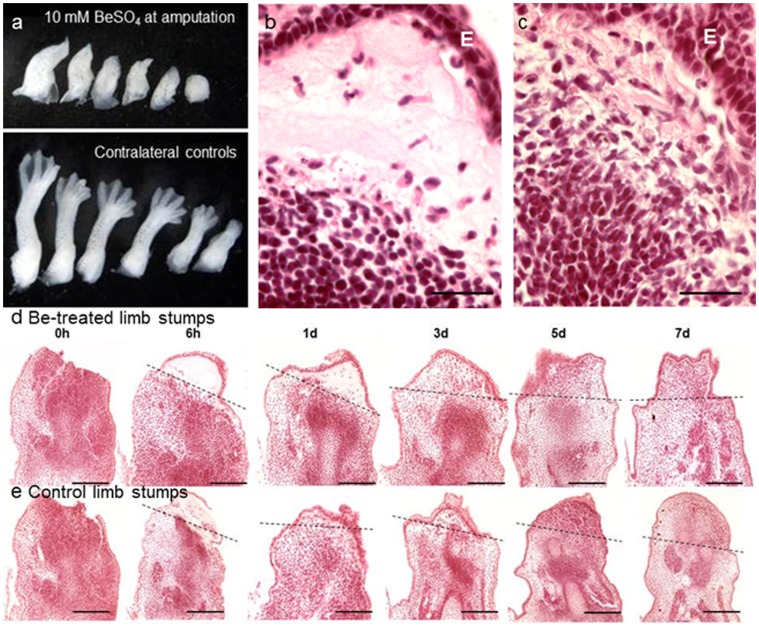
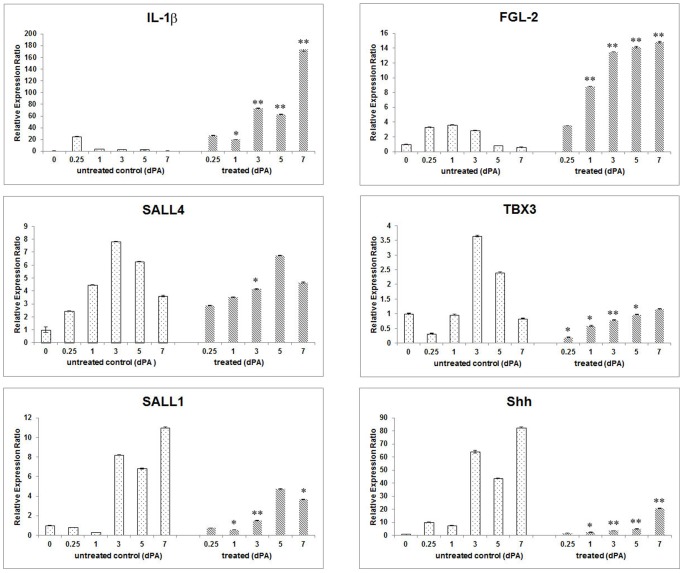
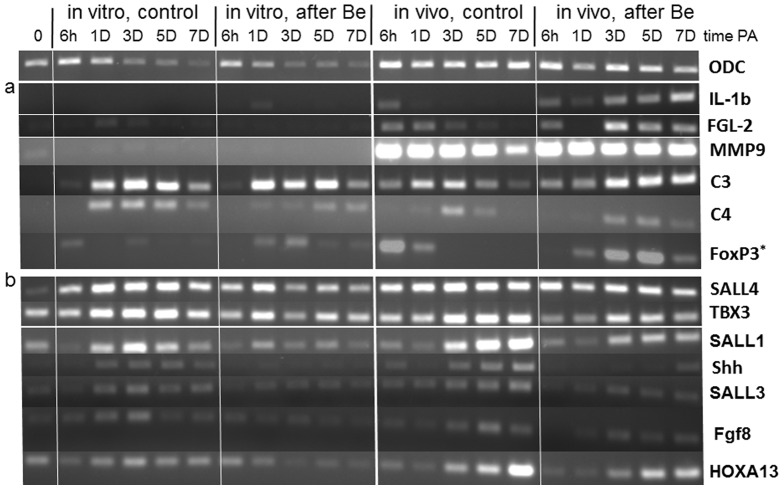
Tissue and organ regeneration, unlike development, involves an injury that in postembryonic animals triggers inflammation followed by resolution. How inflammation affects epimorphic regeneration is largely uninvestigated. Here we examine inflammation and its resolution in Xenopus laevis hindlimb regeneration, which declines during larval development. During the first 5 days postamputation, both regeneration-competent stage 53 and regeneration-deficient stage 57 hindlimbs showed very rapid accumulation of leukocytes and cells expressing interleukin-1β and matrix metalloproteinase 9. Expression of genes for factors mediating inflammatory resolution appeared more persistent at stages 55 and 57 than at stage 53, suggesting changes in this process during development. FoxP3, a marker for regulatory T cells, was upregulated by amputation in limbs at all three stages but only persisted at stage 57, when it was also detected before amputation. Expression of genes for cellular reprogramming, such as SALL4, was upregulated in limbs at all 3 stages, but markers of limb patterning, such as Shh, were expressed later and less actively after amputation in regeneration-deficient limbs. Topical application of specific proinflammatory agents to freshly amputated limbs increased interleukin-1β expression locally. With aqueous solutions of the proinflammatory metal beryllium sulfate, this effect persisted through 7 days postamputation and was accompanied by inhibition of regeneration. In BeSO4-treated limbs expression of markers for both inflammation and resolution, including FoxP3, was prolonged, while genes for cellular reprogramming were relatively unaffected and those for limb patterning failed to be expressed normally. These data imply that in Xenopus hindlimbs postamputation inflammation and its resolution change during development, with little effect on cellular dedifferentiation or reprogramming, but potentially interfering with the expression of genes required for blastema patterning. The results suggest that developmental changes in the larval anuran immune system may be involved in the ontogenetic loss of epimorphic regeneration in this system.
Conflict of interest statement
Figures
References
-
- Dent JN (1962) Limb regeneration in larvae and metamorphosing individuals of the South African clawed toad. J Morphol 110: 61–78. - PubMed
-
- Wolfe AD, Nye HLD, Cameron JA (2000) Extent of ossification at the amputation plane is correlated with the decline of blastema formation and regeneration in Xenopus laevis hindlimbs. Dev Dyn 218: 681–697. - PubMed
-
- Beck CW, Christen B, Slack JM (2003) Molecular pathways needed for regeneration of spinal cord and muscle in a vertebrate. Dev Cell 5: 429–439. - PubMed
-
- Franchini A, Bertolotti E (2011) Tail regenerative capacity and iNOS immunolocalization in Xenopus laevis tadpoles. Cell Tissue Res 344: 261–269. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
