

Proteome analysis and conditional deletion of the EAAT2 glutamate transporter provide evidence against a role of EAAT2 in pancreatic insulin secretion in mice
- PMID: 24280215
- PMCID: PMC3894318
- DOI: 10.1074/jbc.M113.529065
Proteome analysis and conditional deletion of the EAAT2 glutamate transporter provide evidence against a role of EAAT2 in pancreatic insulin secretion in mice
Abstract
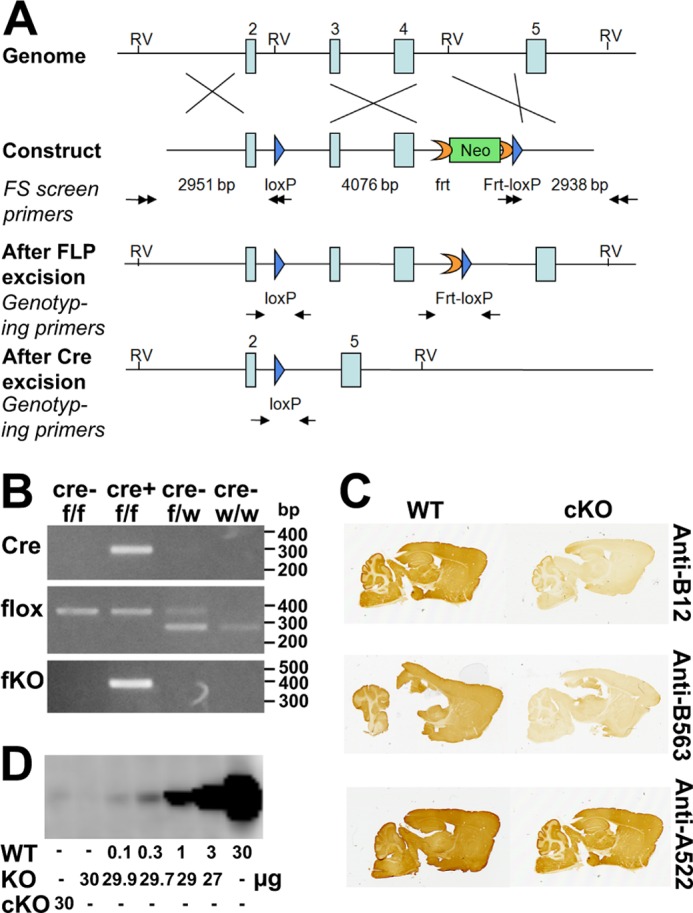
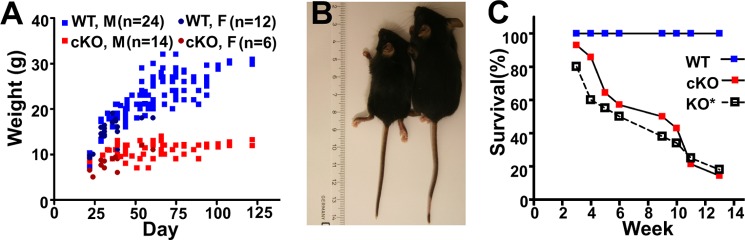
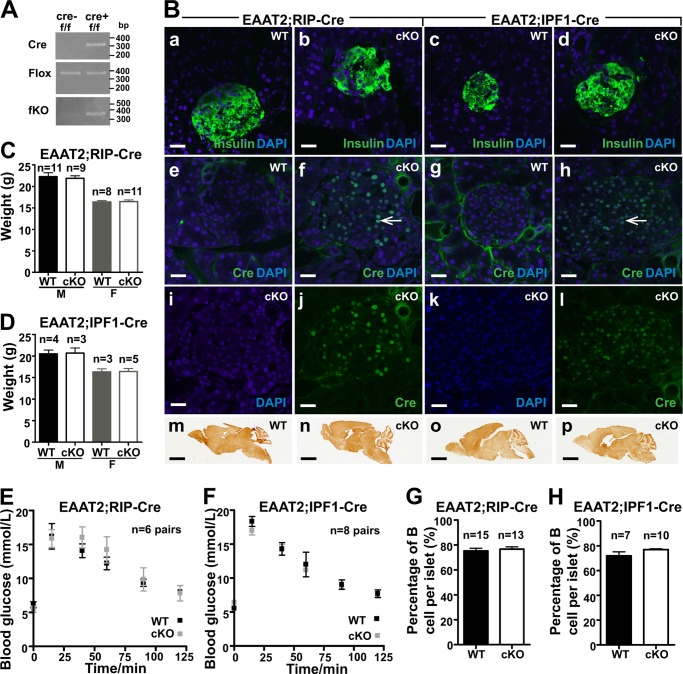
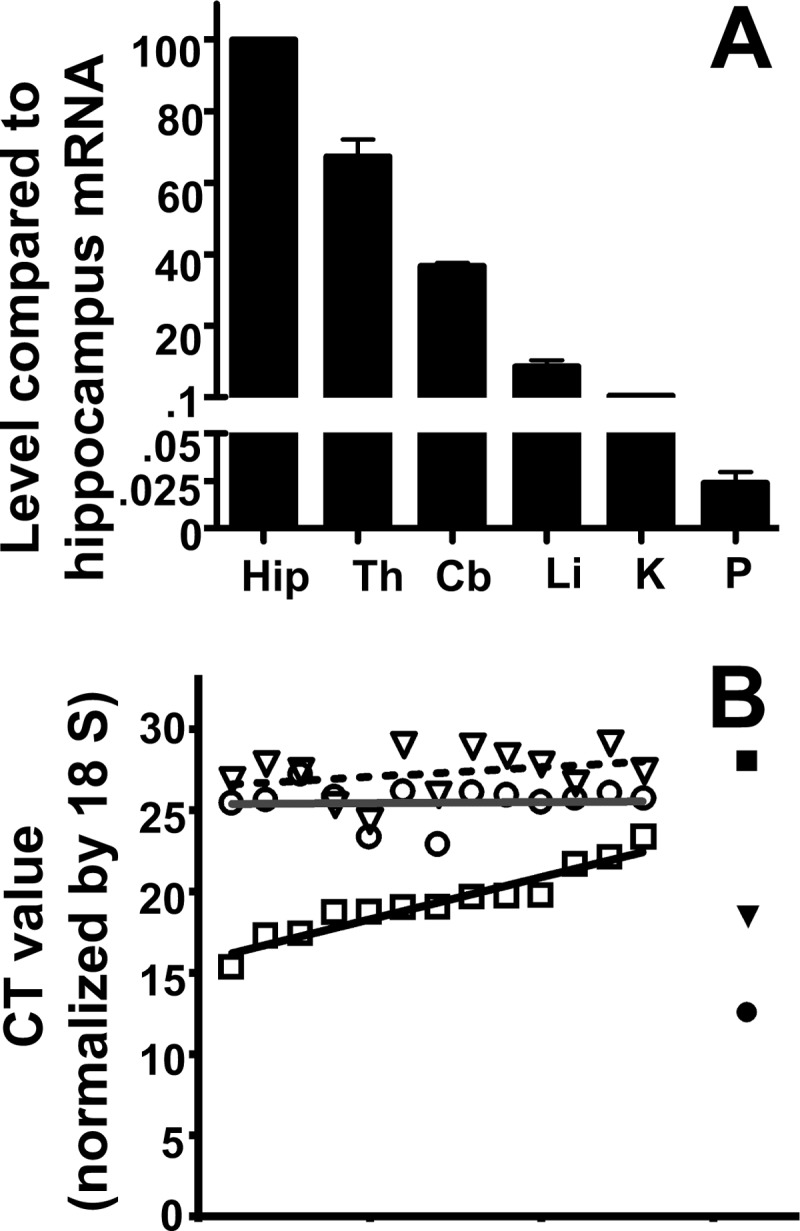
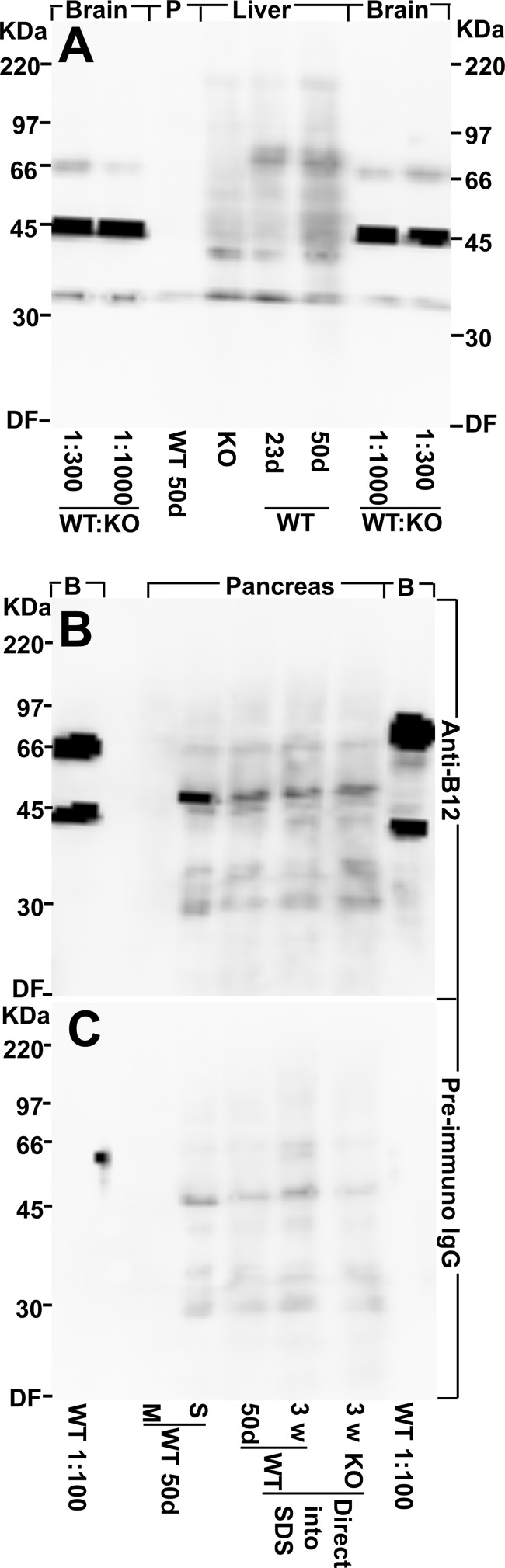
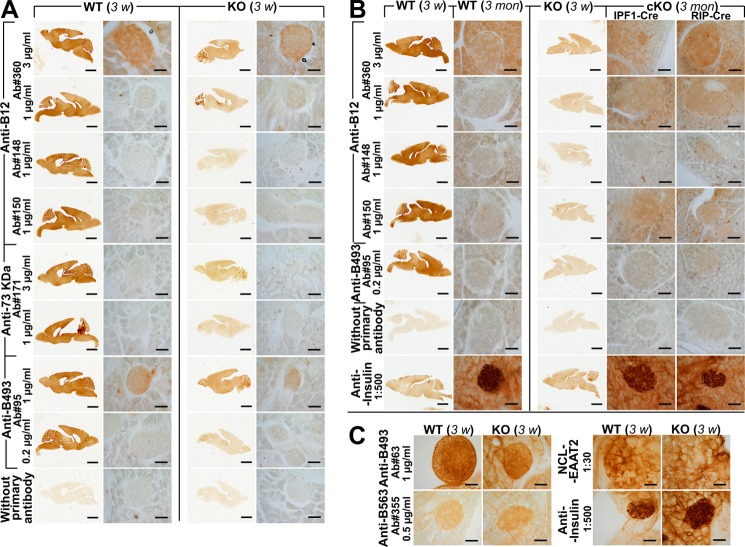
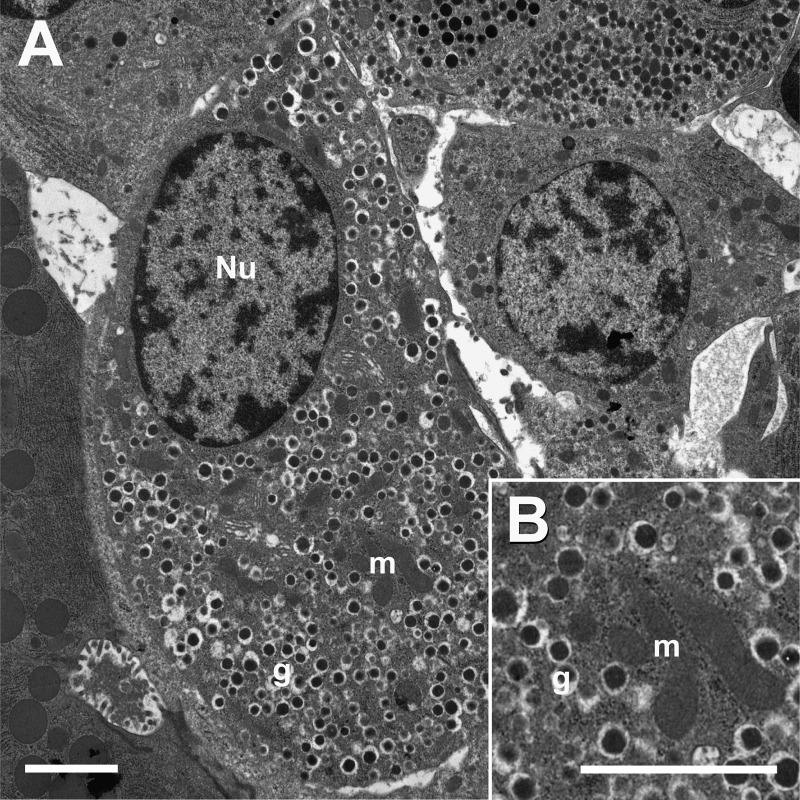
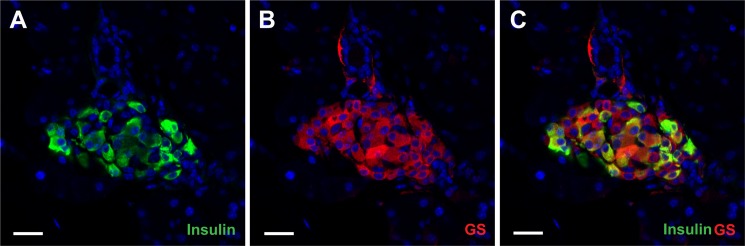
Islet function is incompletely understood in part because key steps in glutamate handling remain undetermined. The glutamate (excitatory amino acid) transporter 2 (EAAT2; Slc1a2) has been hypothesized to (a) provide islet cells with glutamate, (b) protect islet cells against high extracellular glutamate concentrations, (c) mediate glutamate release, or (d) control the pH inside insulin secretory granules. Here we floxed the EAAT2 gene to produce the first conditional EAAT2 knock-out mice. Crossing with Nestin-cyclization recombinase (Cre) eliminated EAAT2 from the brain, resulting in epilepsy and premature death, confirming the importance of EAAT2 for brain function and validating the genetic construction. Crossing with insulin-Cre lines (RIP-Cre and IPF1-Cre) to obtain pancreas-selective deletion did not appear to affect survival, growth, glucose tolerance, or β-cell number. We found (using TaqMan RT-PCR, immunoblotting, immunocytochemistry, and proteome analysis) that the EAAT2 levels were too low to support any of the four hypothesized functions. The proteome analysis detected more than 7,000 islet proteins of which more than 100 were transporters. Although mitochondrial glutamate transporters and transporters for neutral amino acids were present at high levels, all other transporters with known ability to transport glutamate were strikingly absent. Glutamate-metabolizing enzymes were abundant. The level of glutamine synthetase was 2 orders of magnitude higher than that of glutaminase. Taken together this suggests that the uptake of glutamate by islets from the extracellular fluid is insignificant and that glutamate is intracellularly produced. Glutamine synthetase may be more important for islets than assumed previously.
Keywords: Excitatory Amino Acid Transporter; Glutamate Uptake; Glutamine Synthase; Immunohistochemistry; Insulin Secretion; Pancreatic Islets; Proteomics; Slc1a2; Transgenic Mice; VGLUT3.
Figures

Similar articles
-
A role for glutamate transporters in the regulation of insulin secretion.PLoS One. 2011;6(8):e22960. doi: 10.1371/journal.pone.0022960. Epub 2011 Aug 11. PLoS One. 2011. PMID: 21853059 Free PMC article.
-
Hyperglycemia impairs EAAT2 glutamate transporter trafficking and glutamate clearance in islets of Langerhans: implications for type 2 diabetes pathogenesis and treatment.Am J Physiol Endocrinol Metab. 2024 Jul 1;327(1):E27-E41. doi: 10.1152/ajpendo.00069.2024. Epub 2024 May 1. Am J Physiol Endocrinol Metab. 2024. PMID: 38690938 Free PMC article.
-
Axon-terminals expressing EAAT2 (GLT-1; Slc1a2) are common in the forebrain and not limited to the hippocampus.Neurochem Int. 2019 Feb;123:101-113. doi: 10.1016/j.neuint.2018.03.006. Epub 2018 Mar 9. Neurochem Int. 2019. PMID: 29530756
-
Role of excitatory amino acid transporter-2 (EAAT2) and glutamate in neurodegeneration: opportunities for developing novel therapeutics.J Cell Physiol. 2011 Oct;226(10):2484-93. doi: 10.1002/jcp.22609. J Cell Physiol. 2011. PMID: 21792905 Free PMC article. Review.
-
Role of glutamate excitotoxicity and glutamate transporter EAAT2 in epilepsy: Opportunities for novel therapeutics development.Biochem Pharmacol. 2021 Nov;193:114786. doi: 10.1016/j.bcp.2021.114786. Epub 2021 Sep 24. Biochem Pharmacol. 2021. PMID: 34571003 Free PMC article. Review.
Cited by
-
CD44 variant inhibits insulin secretion in pancreatic β cells by attenuating LAT1-mediated amino acid uptake.Sci Rep. 2018 Feb 12;8(1):2785. doi: 10.1038/s41598-018-20973-2. Sci Rep. 2018. PMID: 29434323 Free PMC article.
-
Still in Search for an EAAT Activator: GT949 Does Not Activate EAAT2, nor EAAT3 in Impedance and Radioligand Uptake Assays.ACS Chem Neurosci. 2024 Apr 3;15(7):1424-1431. doi: 10.1021/acschemneuro.3c00731. Epub 2024 Mar 13. ACS Chem Neurosci. 2024. PMID: 38478848 Free PMC article.
-
Multi-omic human pancreatic islet endoplasmic reticulum and cytokine stress response mapping provides type 2 diabetes genetic insights.Cell Metab. 2024 Nov 5;36(11):2468-2488.e7. doi: 10.1016/j.cmet.2024.09.006. Epub 2024 Oct 8. Cell Metab. 2024. PMID: 39383866
-
The betaine/GABA transporter and betaine: roles in brain, kidney, and liver.Front Physiol. 2014 Apr 24;5:159. doi: 10.3389/fphys.2014.00159. eCollection 2014. Front Physiol. 2014. PMID: 24795654 Free PMC article. Review.
-
Factors Influencing Mitochondrial Function as a Key Mediator of Glucose-Induced Insulin Release: Highlighting Nicotinamide Nucleotide Transhydrogenase.Int J Mol Cell Med. 2020 Spring;9(2):107-122. doi: 10.22088/IJMCM.BUMS.9.2.107. Epub 2020 Aug 10. Int J Mol Cell Med. 2020. PMID: 32934948 Free PMC article. Review.
References
-
- Moriyama Y., Hayashi M. (2003) Glutamate-mediated signaling in the islets of Langerhans: a thread entangled. Trends Pharmacol. Sci. 24, 511–517 - PubMed
-
- MacDonald M. J., Fahien L. A. (2000) Glutamate is not a messenger in insulin secretion. J. Biol. Chem. 275, 34025–34027 - PubMed
-
- Yamada S., Komatsu M., Sato Y., Yamauchi K., Aizawa T., Hashizume K. (2001) Glutamate is not a major conveyer of ATP-sensitive K+ channel-independent glucose action in pancreatic islet β cell. Endocr. J. 48, 391–395 - PubMed
-
- Bertrand G., Ishiyama N., Nenquin M., Ravier M. A., Henquin J.-C. (2002) The elevation of glutamate content and the amplification of insulin secretion in glucose-stimulated pancreatic islets are not causally related. J. Biol. Chem. 277, 32883–32891 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
