

A novel mercuric reductase from the unique deep brine environment of Atlantis II in the Red Sea
- PMID: 24280218
- PMCID: PMC3894346
- DOI: 10.1074/jbc.M113.493429
A novel mercuric reductase from the unique deep brine environment of Atlantis II in the Red Sea
Abstract
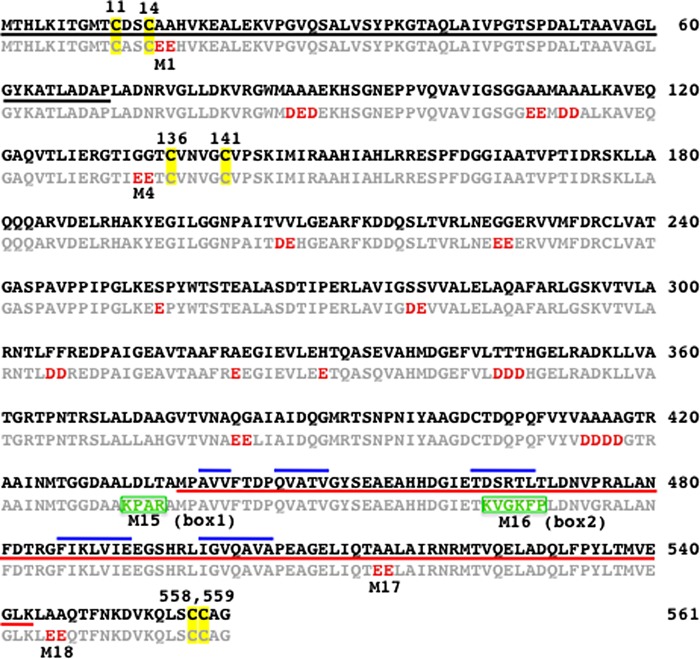
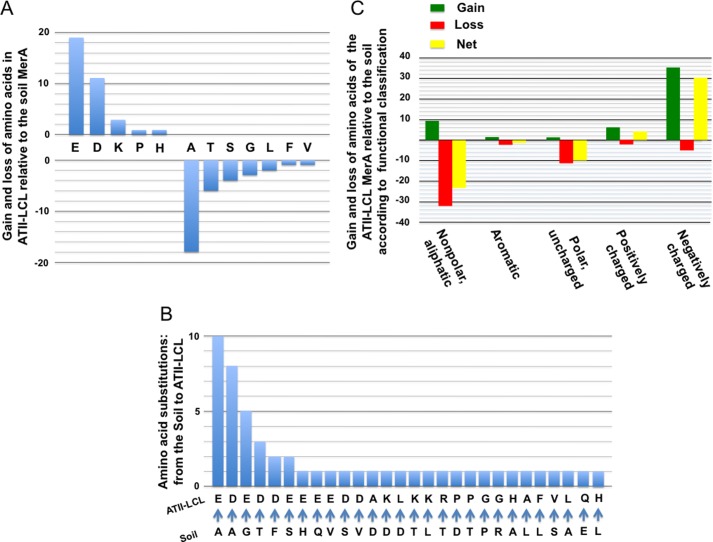
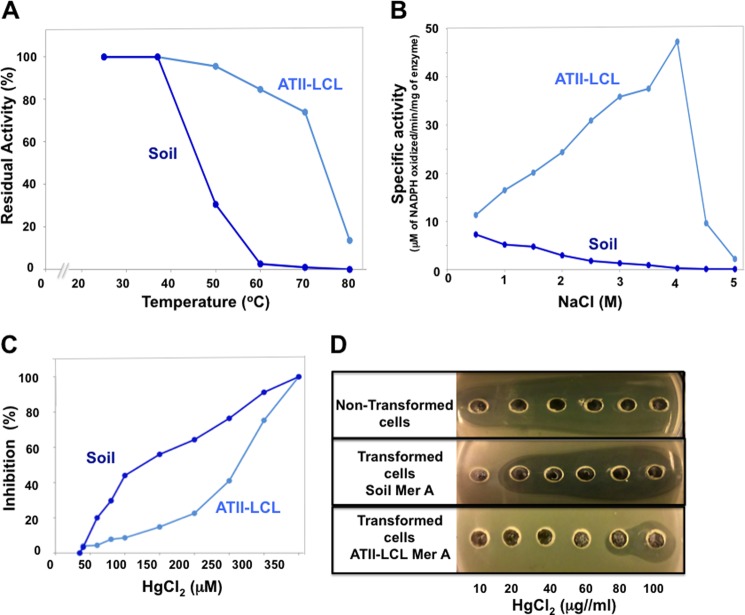
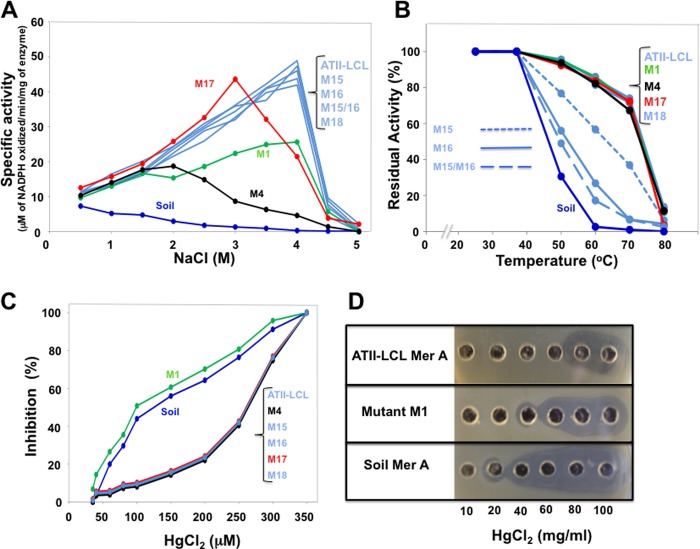
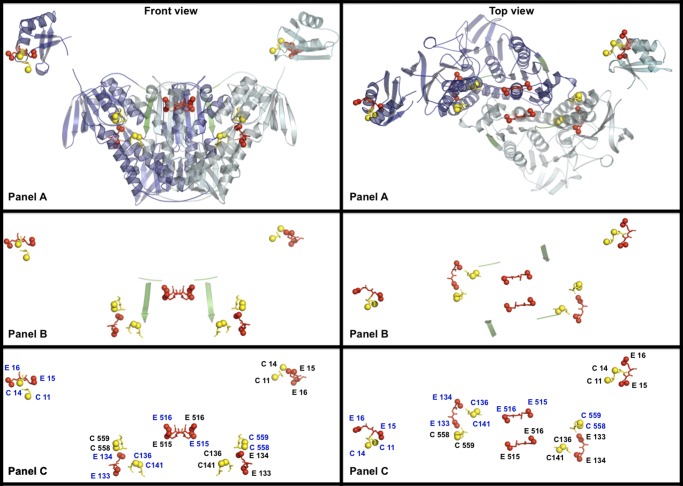
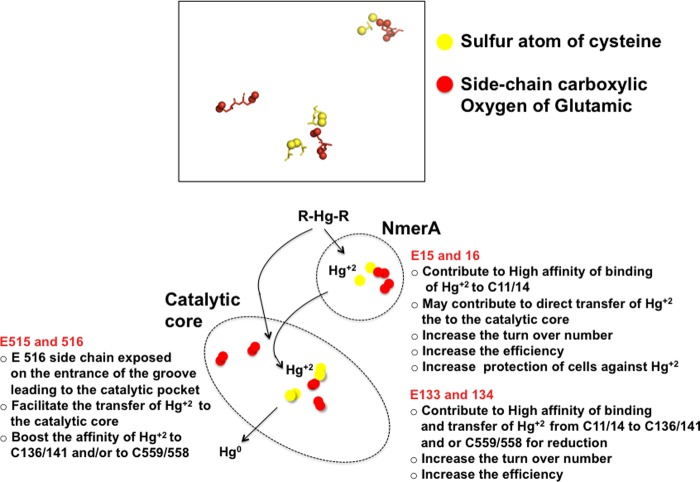
A unique combination of physicochemical conditions prevails in the lower convective layer (LCL) of the brine pool at Atlantis II (ATII) Deep in the Red Sea. With a maximum depth of over 2000 m, the pool is characterized by acidic pH (5.3), high temperature (68 °C), salinity (26%), low light levels, anoxia, and high concentrations of heavy metals. We have established a metagenomic dataset derived from the microbial community in the LCL, and here we describe a gene for a novel mercuric reductase, a key component of the bacterial detoxification system for mercuric and organomercurial species. The metagenome-derived gene and an ortholog from an uncultured soil bacterium were synthesized and expressed in Escherichia coli. The properties of their products show that, in contrast to the soil enzyme, the ATII-LCL mercuric reductase is functional in high salt, stable at high temperatures, resistant to high concentrations of Hg(2+), and efficiently detoxifies Hg(2+) in vivo. Interestingly, despite the marked functional differences between the orthologs, their amino acid sequences differ by less than 10%. Site-directed mutagenesis and kinetic analysis of the mutant enzymes, in conjunction with three-dimensional modeling, have identified distinct structural features that contribute to extreme halophilicity, thermostability, and high detoxification capacity, suggesting that these were acquired independently during the evolution of this enzyme. Thus, our work provides fundamental structural insights into a novel protein that has undergone multiple biochemical and biophysical adaptations to promote the survival of microorganisms that reside in the extremely demanding environment of the ATII-LCL.
Keywords: Atlantis II Brine Pool; Enzyme Kinetics; Enzyme Mechanisms; Enzyme Structure; Extreme Halophilic; Mercuric Reductase; Metagenomics; Mutagenesis Site-specific; Red Sea Atlantis II; Thermophilic.
Figures
Similar articles
-
Thermal Stability of a Mercuric Reductase from the Red Sea Atlantis II Hot Brine Environment as Analyzed by Site-Directed Mutagenesis.Appl Environ Microbiol. 2019 Jan 23;85(3):e02387-18. doi: 10.1128/AEM.02387-18. Print 2019 Feb 1. Appl Environ Microbiol. 2019. PMID: 30446558 Free PMC article.
-
Molecular Adaptations of Bacterial Mercuric Reductase to the Hypersaline Kebrit Deep in the Red Sea.Appl Environ Microbiol. 2019 Feb 6;85(4):e01431-18. doi: 10.1128/AEM.01431-18. Print 2019 Feb 15. Appl Environ Microbiol. 2019. PMID: 30504211 Free PMC article.
-
Red Sea Atlantis II brine pool nitrilase with unique thermostability profile and heavy metal tolerance.BMC Biotechnol. 2016 Feb 11;16:14. doi: 10.1186/s12896-016-0244-2. BMC Biotechnol. 2016. PMID: 26868129 Free PMC article.
-
Bacterial resistances to inorganic mercury salts and organomercurials.Plasmid. 1992 Jan;27(1):4-16. doi: 10.1016/0147-619x(92)90002-r. Plasmid. 1992. PMID: 1311113 Review.
-
Prokaryotic lifestyles in deep sea habitats.Extremophiles. 2008 Jan;12(1):15-25. doi: 10.1007/s00792-006-0059-5. Epub 2007 Jan 17. Extremophiles. 2008. PMID: 17225926 Review.
Cited by
-
Biochemical and Structural Properties of a Thermostable Mercuric Ion Reductase from Metallosphaera sedula.Front Bioeng Biotechnol. 2015 Jul 13;3:97. doi: 10.3389/fbioe.2015.00097. eCollection 2015. Front Bioeng Biotechnol. 2015. PMID: 26217660 Free PMC article.
-
Toxicomicrobiomics: The Human Microbiome vs. Pharmaceutical, Dietary, and Environmental Xenobiotics.Front Pharmacol. 2020 Apr 16;11:390. doi: 10.3389/fphar.2020.00390. eCollection 2020. Front Pharmacol. 2020. PMID: 32372951 Free PMC article. Review.
-
Bioprospecting of Novel Extremozymes From Prokaryotes-The Advent of Culture-Independent Methods.Front Microbiol. 2021 Feb 10;12:630013. doi: 10.3389/fmicb.2021.630013. eCollection 2021. Front Microbiol. 2021. PMID: 33643258 Free PMC article. Review.
-
Novel Enzymes From the Red Sea Brine Pools: Current State and Potential.Front Microbiol. 2021 Oct 27;12:732856. doi: 10.3389/fmicb.2021.732856. eCollection 2021. Front Microbiol. 2021. PMID: 34777282 Free PMC article. Review.
-
Deep Hypersaline Anoxic Basins as Untapped Reservoir of Polyextremophilic Prokaryotes of Biotechnological Interest.Mar Drugs. 2020 Jan 30;18(2):91. doi: 10.3390/md18020091. Mar Drugs. 2020. PMID: 32019162 Free PMC article. Review.
References
-
- Backer H., Schoell M. (1972) New deeps with brines and metalliferous sediments in the Red Sea. Nat. Phys. Sci. 240, 153–158
-
- Miller A. R., Densmore C. D., Degens E. T., Hathaway J. C., Manheim F. T., McFarlin P. J., Pocklington R., Jokelin A. (1966) Hot brines and recent iron deposits in deeps of the Red Sea. Geochim. Cosmochim. Acta 30, 341–360
-
- Swift S. A., Bower A. S., Schmitt R. W. (2012) Vertical, horizontal, and temporal changes in temperature in the Atlantis II and Discovery hot brine pools, Red Sea. Deep-Sea Res. I 64, 118–128
-
- Antunes A., Ngugi D. K., Stingl U. (2011) Microbiology of the Red Sea (and other) deep-sea anoxic brine lakes. Environ. Microbiol. Rep. 3, 416–433 - PubMed
-
- Barkay T., Wagner-Döbler I. (2005) Microbial transformations of mercury: potentials, challenges, and achievements in controlling mercury toxicity in the environment. Adv. Appl. Microbiol. 57, 1–52 - PubMed
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
