

RNAi mediated down regulation of myo-inositol-3-phosphate synthase to generate low phytate rice
- PMID: 24280240
- PMCID: PMC4883737
- DOI: 10.1186/1939-8433-6-12
RNAi mediated down regulation of myo-inositol-3-phosphate synthase to generate low phytate rice
Abstract
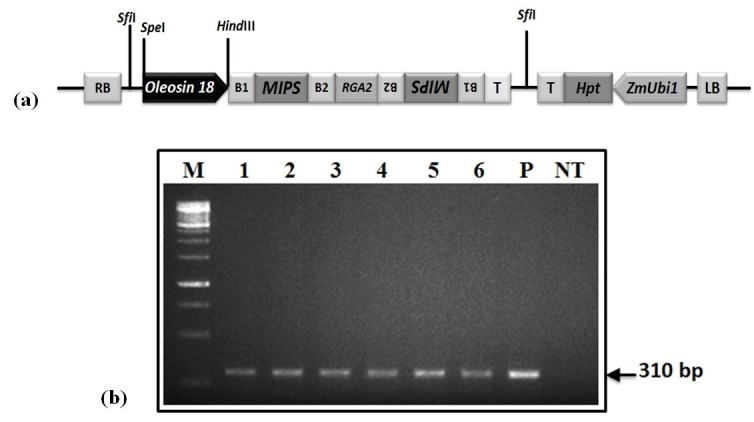
Background: Phytic acid (InsP6) is considered as the major source of phosphorus and inositol phosphates in cereal grains. Reduction of phytic acid level in cereal grains is desirable in view of its antinutrient properties to maximize mineral bioavailability and minimize the load of phosphorus waste management. We report here RNAi mediated seed-specific silencing of myo-inositol-3-phosphate synthase (MIPS) gene catalyzing the first step of phytic acid biosynthesis in rice. Moreover, we also studied the possible implications of MIPS silencing on myo-inositol and related metabolism, since, first step of phytic acid biosynthesis is also the rate limiting step of myo-inositol synthesis, catalyzed by MIPS.
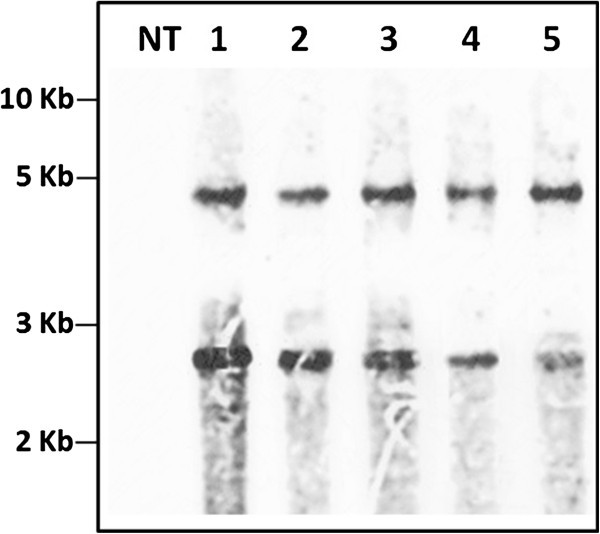
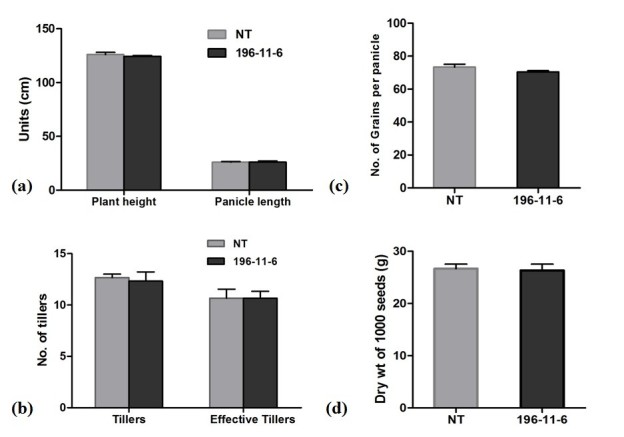
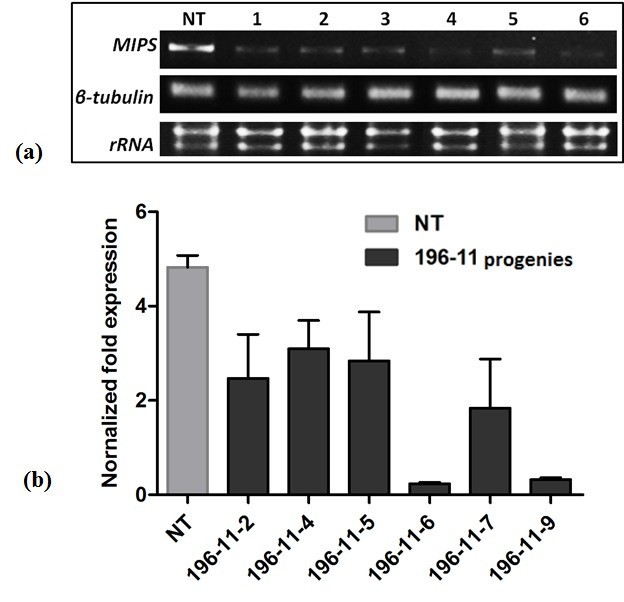
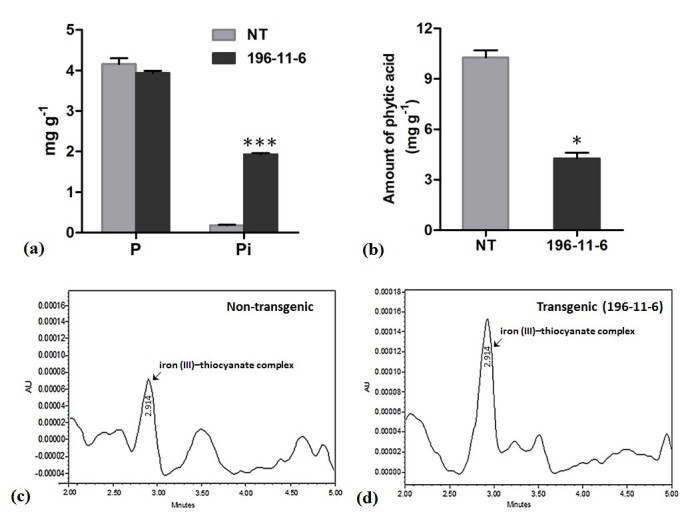
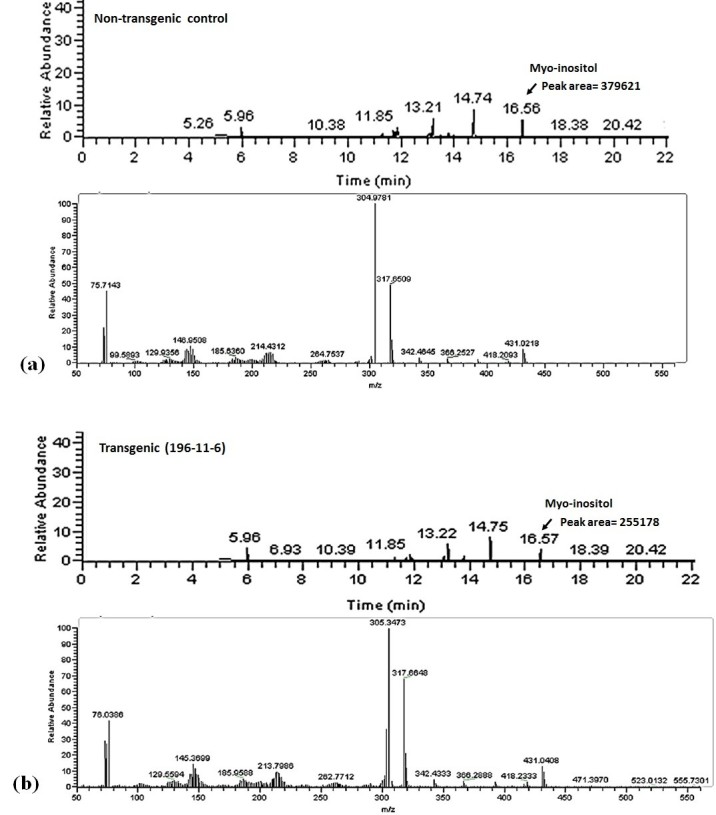
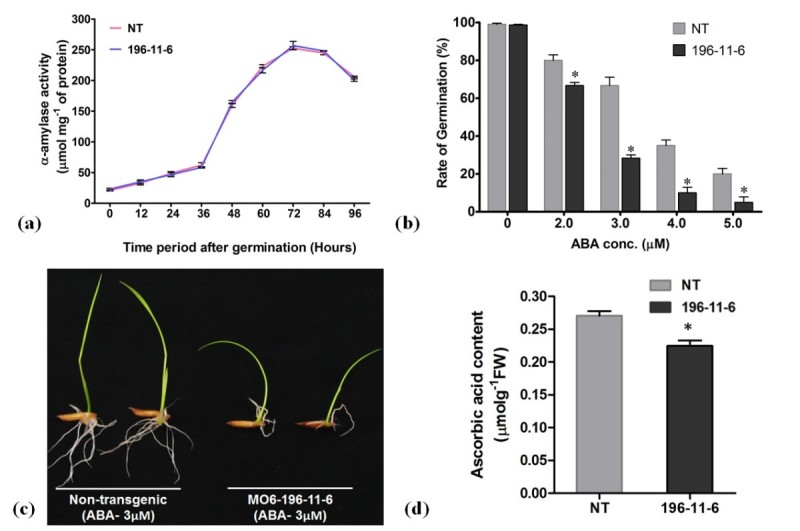
Results: The resulting transgenic rice plants (T3) showed a 4.59 fold down regulation in MIPS gene expression, which corresponds to a significant decrease in phytate levels and a simultaneous increment in the amount of inorganic phosphate in the seeds. A diminution in the myo-inositol content of transgenic plants was also observed due to disruption of the first step of phytic acid biosynthetic pathway, which further reduced the level of ascorbate and altered abscisic acid (ABA) sensitivity of the transgenic plants. In addition, our results shows that in the transgenic plants, the lower phytate levels has led to an increment of divalent cations, of which a 1.6 fold increase in the iron concentration in milled rice seeds was noteworthy. This increase could be attributed to reduced chelation of divalent metal (iron) cations, which may correlate to higher iron bioavailability in the endosperm of rice grains.
Conclusion: The present study evidently suggests that seed-specific silencing of MIPS in transgenic rice plants can yield substantial reduction in levels of phytic acid along with an increase in inorganic phosphate content. However, it was also demonstrated that the low phytate seeds had an undesirable diminution in levels of myo-inositol and ascorbate, which probably led to sensitiveness of seeds to abscisic acid during germination. Therefore, it is suggested that though MIPS is the prime target for generation of low phytate transgenic plants, down-regulation of MIPS can have detrimental effect on myo-inositol synthesis and related pathways which are involved in key plant metabolism.
Figures
References
-
- Alimohammadi M, Silva K, Ballu C, Ali N, Khodakovskaya MV. Reduction of inositol (1,4,5)-trisphosphate affects the overall phosphoinositol pathway and leads to modifications in light signaling and secondary metabolism in tomato plants. J Exp Bot. 2012;63:825–835. doi: 10.1093/jxb/err306. - DOI - PMC - PubMed
-
- Bernfeld P. Amylases α and β. In: Colowick SP, Kalpan NO, editors. Methods in Enzymology. New York 1: Academic Press; 1955. pp. 149–158.
-
- Borg S, Brinch-Pedersen H, Tauris B, Holm PB. Iron transport, deposition and bioavailability in the wheat and barley grain. Plant Soil. 2009;325:15–24. doi: 10.1007/s11104-009-0046-6. - DOI
LinkOut - more resources
Full Text Sources
Other Literature Sources
