

Role of class III phosphatidylinositol 3-kinase during programmed nuclear death of Tetrahymena thermophila
- PMID: 24280724
- PMCID: PMC5396089
- DOI: 10.4161/auto.26929
Role of class III phosphatidylinositol 3-kinase during programmed nuclear death of Tetrahymena thermophila
Abstract
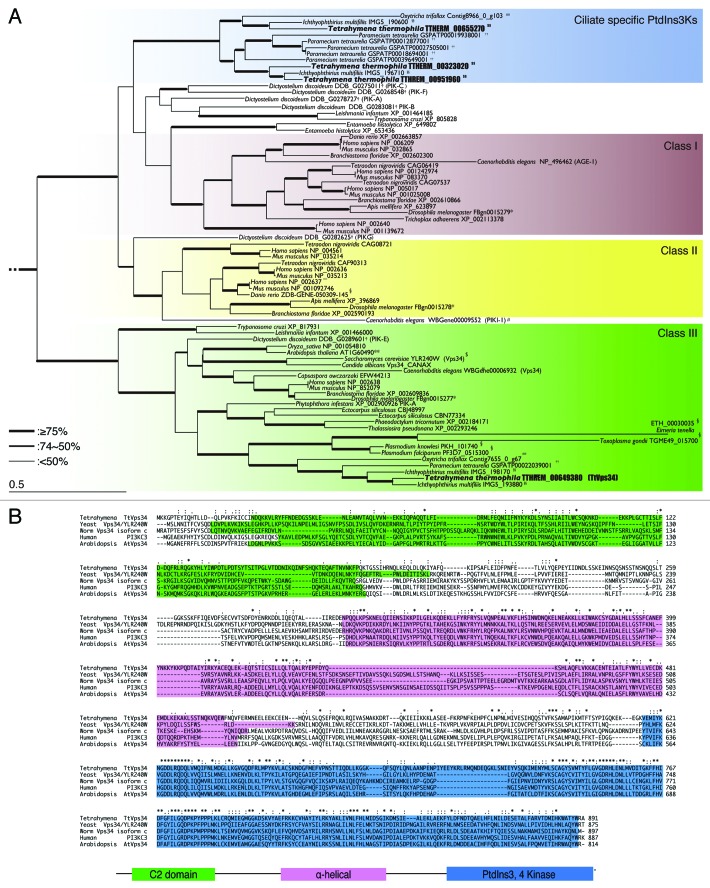
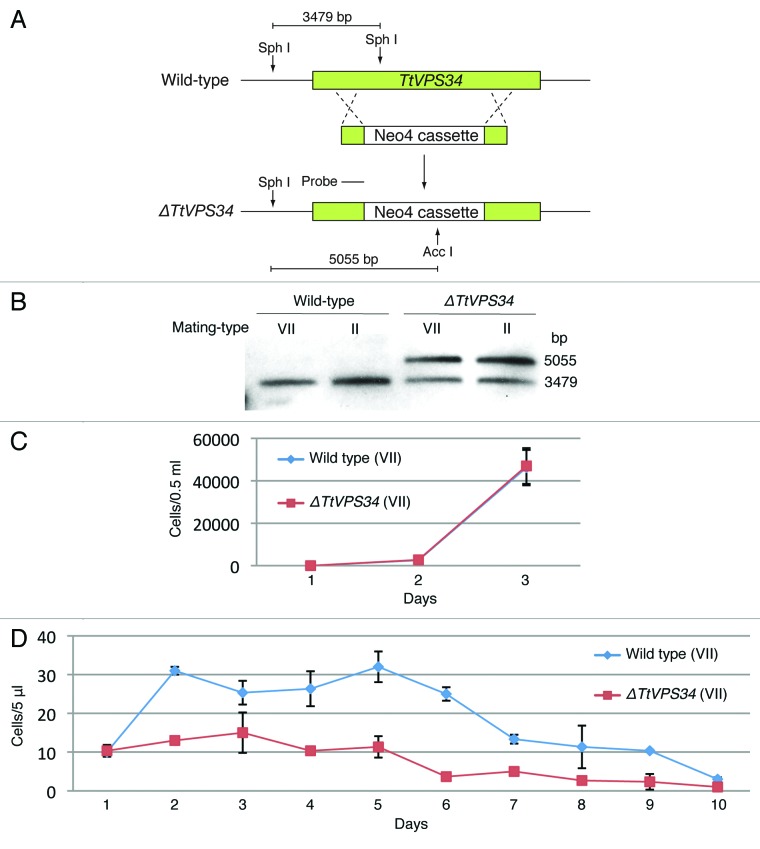
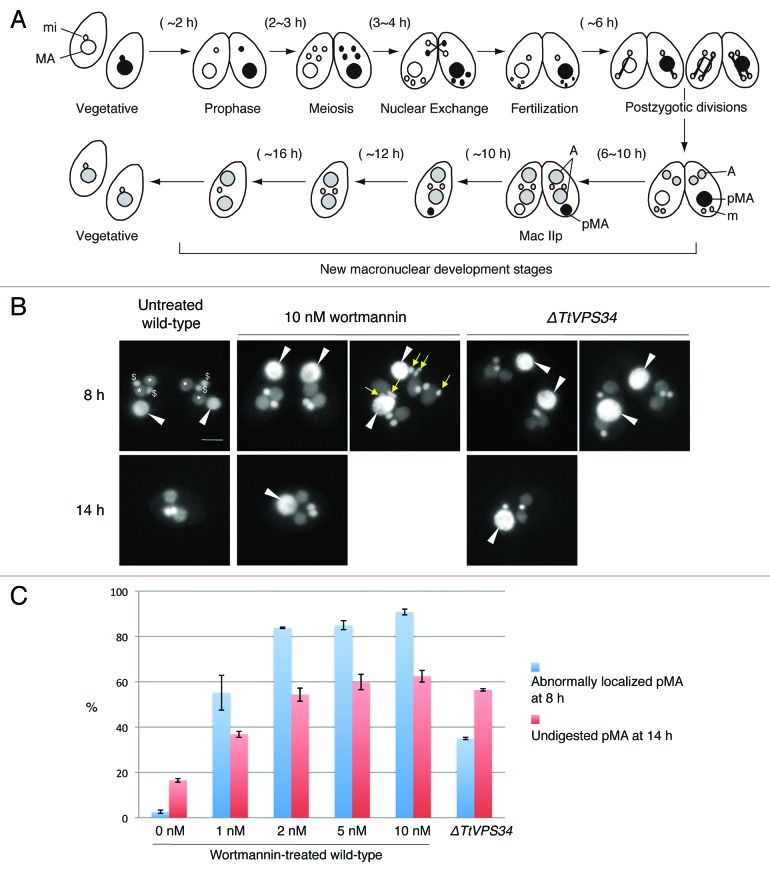
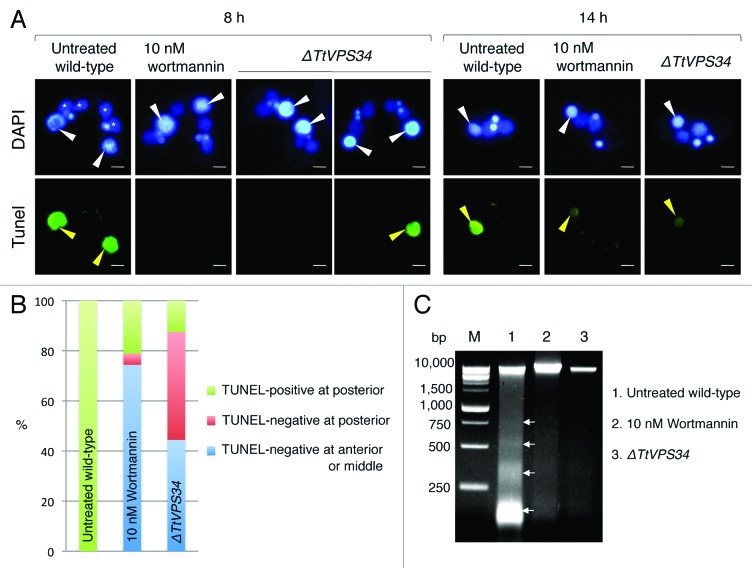
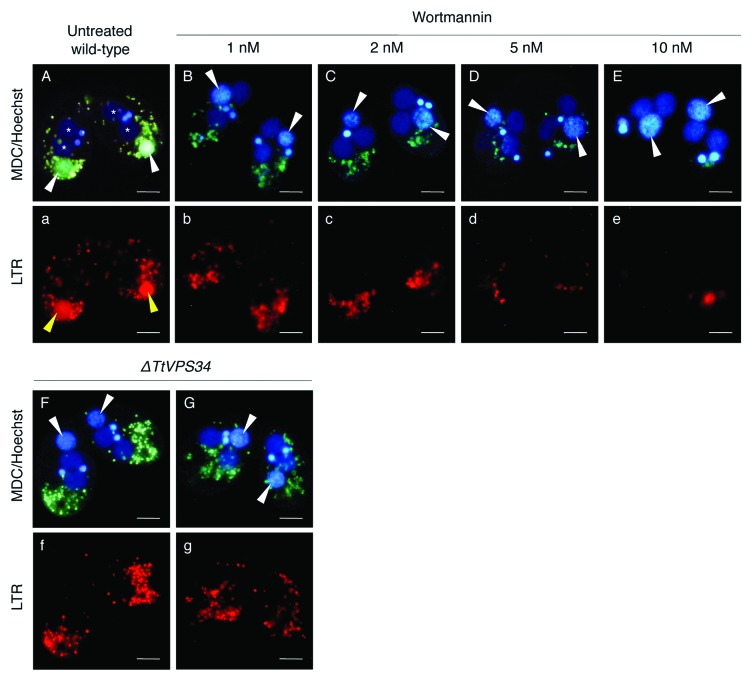
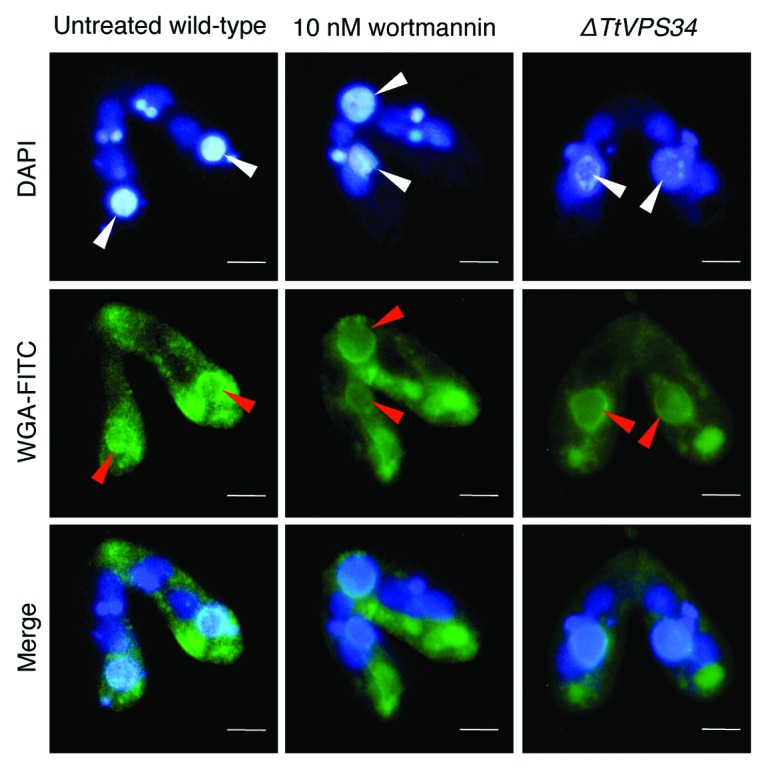
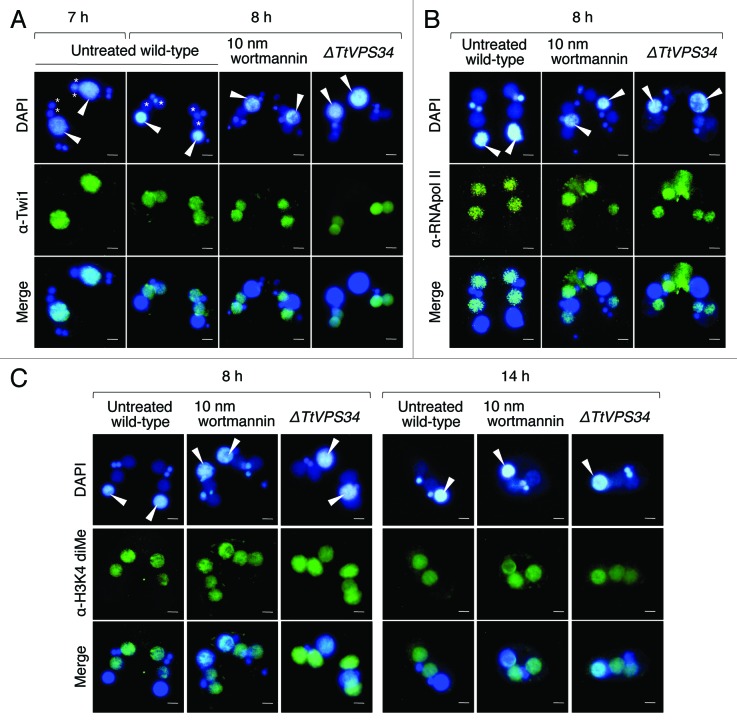
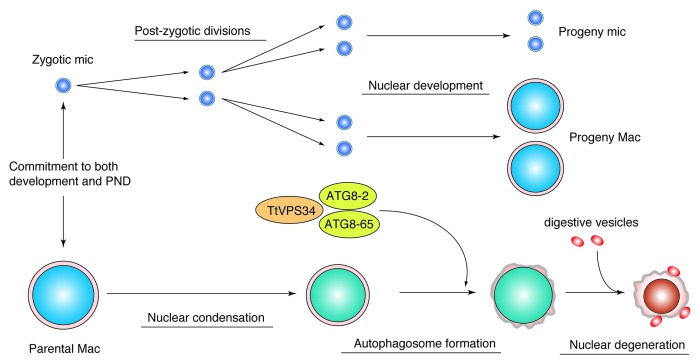
Programmed nuclear death (PND) in the ciliate protozoan Tetrahymena thermophila is a novel type of autophagy that occurs during conjugation, in which only the parental somatic macronucleus is destined to die and is then eliminated from the progeny cytoplasm. Other coexisting nuclei, however, such as new micro- and macronuclei are unaffected. PND starts with condensation in the nucleus followed by apoptotic DNA fragmentation, lysosomal acidification, and final resorption. Because of the peculiarity in the process and the absence of some ATG genes in this organism, the mechanism of PND has remained unclear. In this study, we focus on the role of class III phosphatidylinositol 3-kinase (PtdIns3K, corresponding to yeast Vps34) in order to identify central regulators of PND. We identified the sole Tetrahymena thermophila ortholog (TtVPS34) to yeast Vps34 and human PIK3C3 (the catalytic subunit of PtdIns3K), through phylogenetic analysis, and generated the gene knockdown mutant for functional analysis. Loss of TtVPS34 activity prevents autophagosome formation on the parental macronucleus, and this nucleus escapes from the lysosomal pathway. In turn, DNA fragmentation and final resorption of the nucleus are drastically impaired. These phenotypes are similar to the situation in the ATG8Δ mutants of Tetrahymena, implying an inextricable link between TtVPS34 and TtATG8s in controlling PND as well as general macroautophagy. On the other hand, TtVPS34 does not appear responsible for the nuclear condensation and does not affect the progeny nuclear development. These results demonstrate that TtVPS34 is critically involved in the nuclear degradation events of PND in autophagosome formation rather than with an involvement in commitment to the death program.
Keywords: Atg8; PtdIns3K; Tetrahymena; Vps34; conjugation; macroautophagy; nuclear apoptosis; nuclear development.
Figures
References
-
- Chalker DL. . Transformation and strain engineering of Tetrahymena. . Methods Cell Biol 2012; 109:327 - 45; http://dx.doi.org/ 10.1016/B978-0-12-385967-9.00011-6; PMID: 22444150 - DOI - PubMed
-
- Coyne RS, Stover NA, Miao W. . Whole genome studies of Tetrahymena. . Methods Cell Biol 2012; 109:53 - 81; http://dx.doi.org/ 10.1016/B978-0-12-385967-9.00004-9; PMID: 22444143 - DOI - PubMed
-
- Orias E, Cervantes MD, Hamilton EP. . Tetrahymena thermophila, a unicellular eukaryote with separate germline and somatic genomes. Res Microbiol 2011; 162:578 - 86; http://dx.doi.org/ 10.1016/j.resmic.2011.05.001; PMID: 21624459 - DOI - PMC - PubMed
-
- Iwamoto M, Mori C, Kojidani T, Bunai F, Hori T, Fukagawa T, Hiraoka Y, Haraguchi T. . Two distinct repeat sequences of Nup98 nucleoporins characterize dual nuclei in the binucleated ciliate tetrahymena. . Curr Biol 2009; 19:843 - 7; http://dx.doi.org/ 10.1016/j.cub.2009.03.055; PMID: 19375312 - DOI - PubMed
-
- Cole E, Sugai T. . Developmental progression of Tetrahymena through the cell cycle and conjugation. Methods Cell Biol 2012; 109:177 - 236; http://dx.doi.org/ 10.1016/B978-0-12-385967-9.00007-4; PMID: 22444146 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources