

Triggers of key calcium signals during erythrocyte invasion by Plasmodium falciparum
- PMID: 24280897
- PMCID: PMC3868333
- DOI: 10.1038/ncomms3862
Triggers of key calcium signals during erythrocyte invasion by Plasmodium falciparum
Abstract
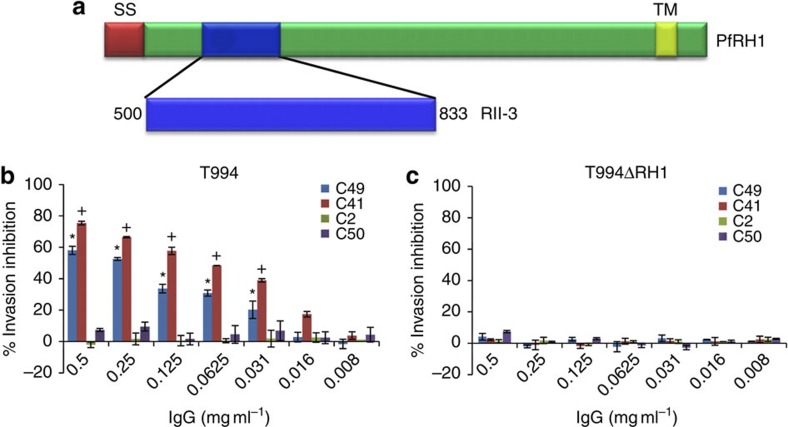
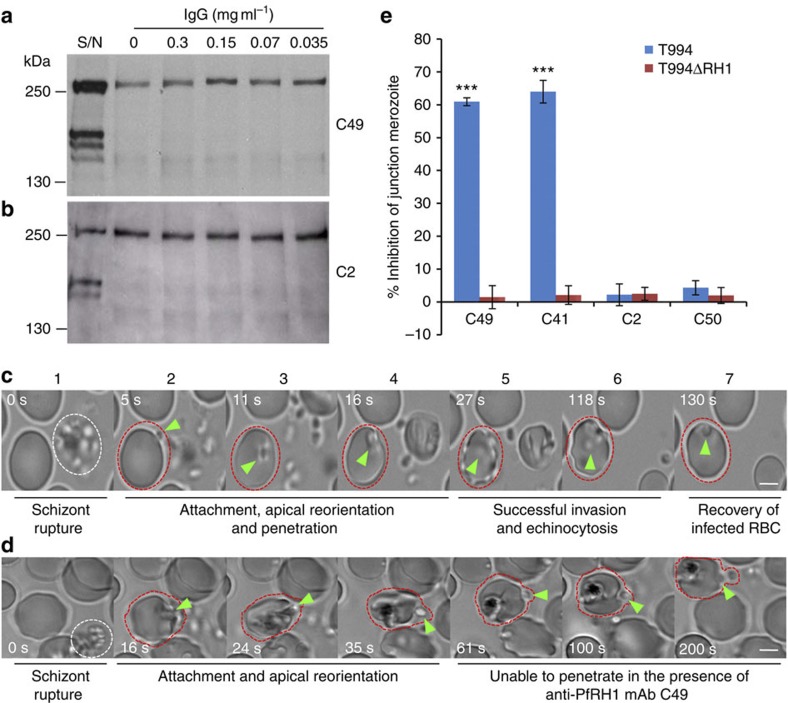
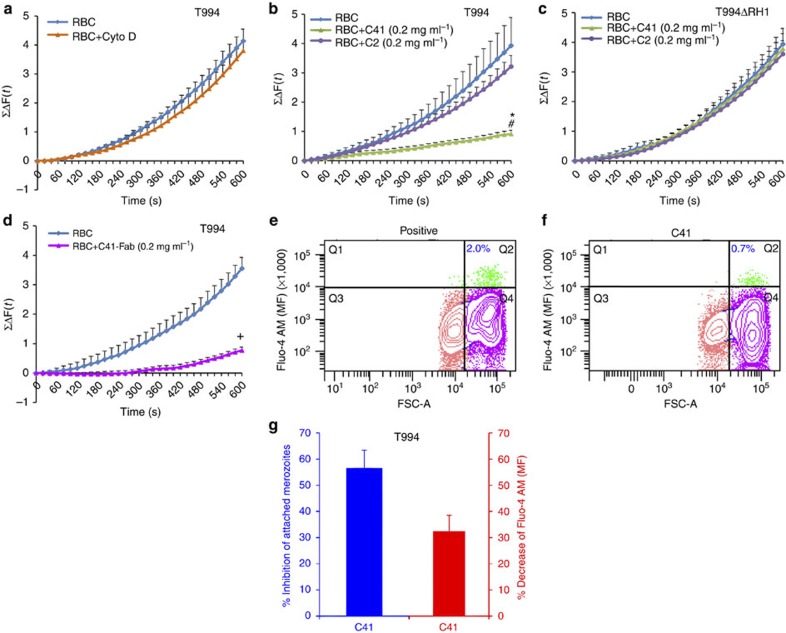
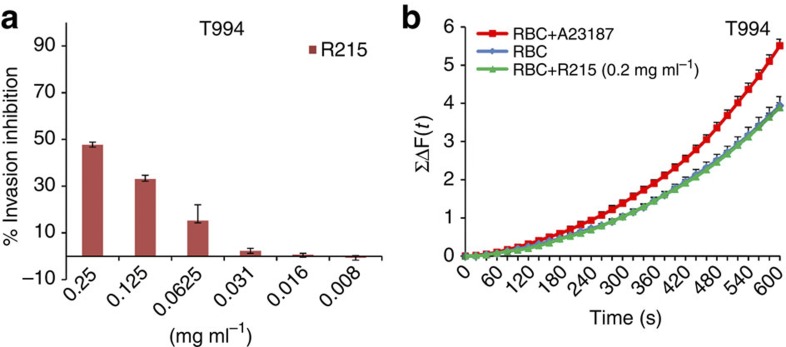
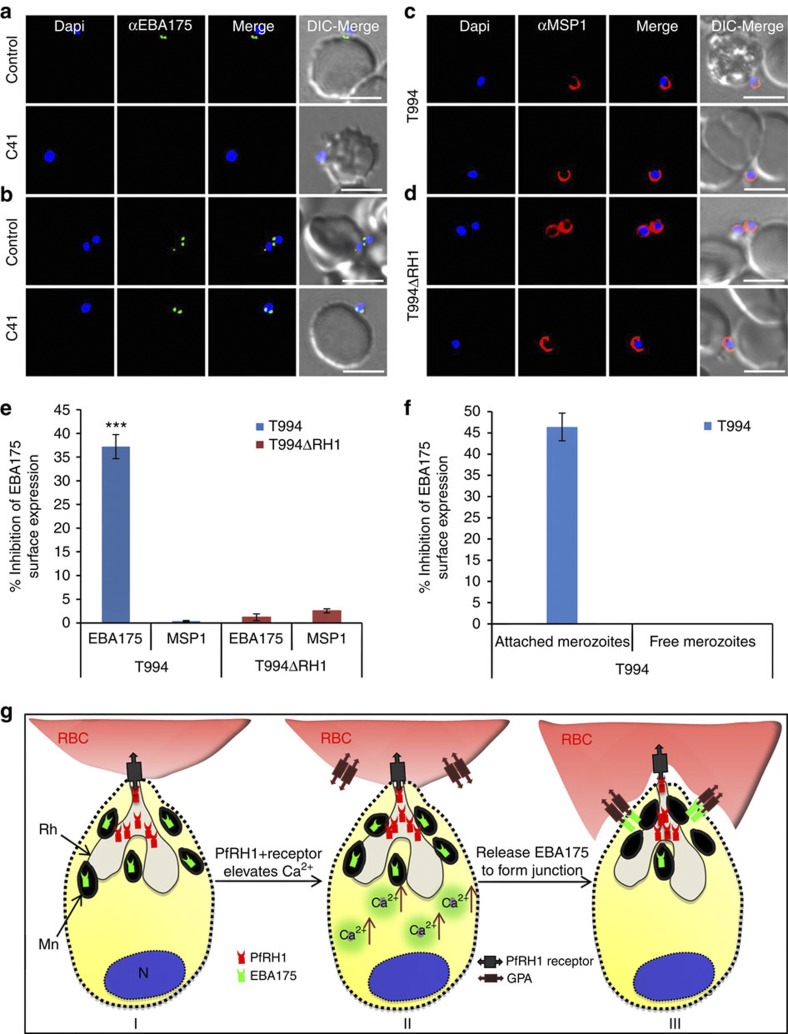
Invasion of erythrocytes by Plasmodium falciparum merozoites is a complex multi-step process mediated by specific interactions between host receptors and parasite ligands. Reticulocyte-binding protein homologues (RHs) and erythrocyte-binding-like (EBL) proteins are discharged from specialized organelles and used in early steps of invasion. Here we show that monoclonal antibodies against PfRH1 (an RH) block merozoite invasion by specifically inhibiting calcium signalling in the parasite, whereas invasion-inhibiting monoclonal antibodies targeting EBA175 (an EBL protein) have no effect on signalling. We further show that inhibition of this calcium signalling prevents EBA175 discharge and thereby formation of the junction between parasite and host cell. Our results indicate that PfRH1 has an initial sensing as well as signal transduction role that leads to the subsequent release of EBA175. They also provide new insights on how RH-host cell interactions lead to essential downstream signalling events in the parasite, suggesting new targets for malaria intervention.
Figures

References
-
- Cowman A. F. & Crabb B. S. Invasion of red blood cells by malaria parasites. Cell 124, 755–766 (2006). - PubMed
-
- Baum J., Gilberger T. W., Frischknecht F. & Meissner M. Host-cell invasion by malaria parasites: insights from Plasmodium and Toxoplasma. Trends Parasitol. 24, 557–563 (2008). - PubMed
-
- Gaur D. & Chitnis C. E. Molecular interactions and signalling mechanisms during erythrocyte invasion by malaria parasites. Curr. Opin. Microbiol. 14, 422–428 (2011). - PubMed
-
- Gunalan K., Gao X., Yap S. S., Huang X. & Preiser P. R. The role of the reticulocyte-binding-like protein homologues of Plasmodium in erythrocyte sensing and invasion. Cell Microbiol. 15, 35–44 (2013). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
