

Pathways of lipid metabolism in marine algae, co-expression network, bottlenecks and candidate genes for enhanced production of EPA and DHA in species of Chromista
- PMID: 24284429
- PMCID: PMC3853752
- DOI: 10.3390/md11114662
Pathways of lipid metabolism in marine algae, co-expression network, bottlenecks and candidate genes for enhanced production of EPA and DHA in species of Chromista
Abstract
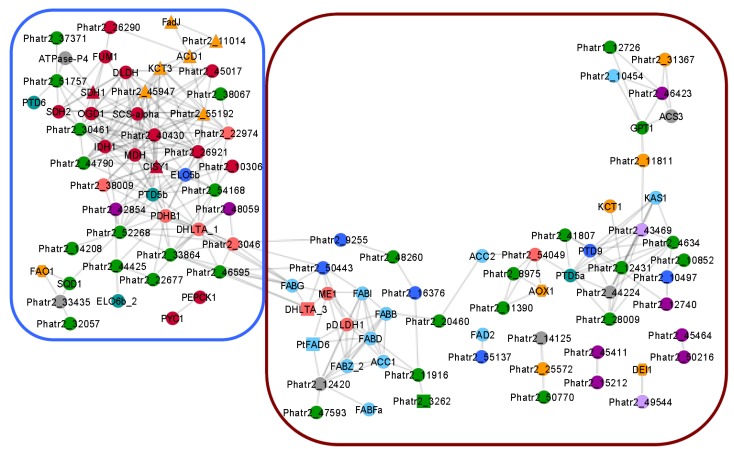
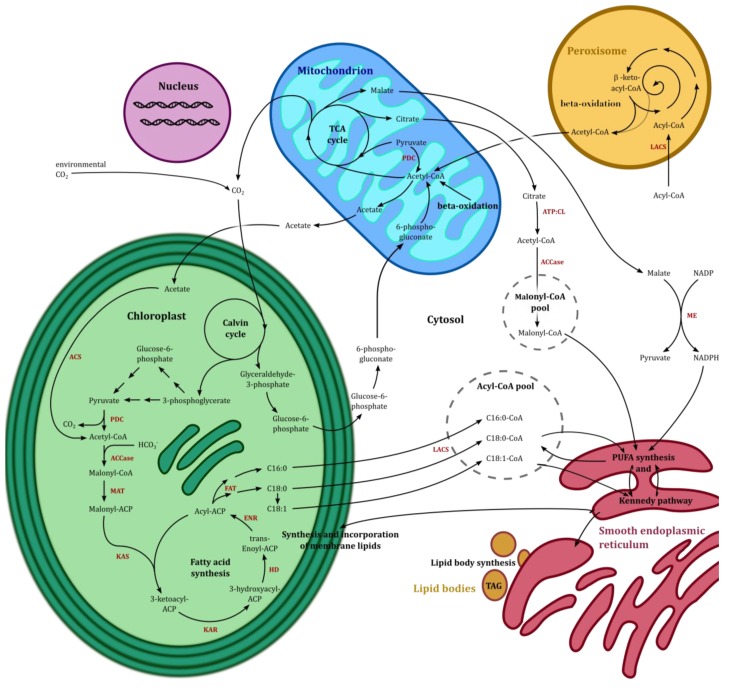
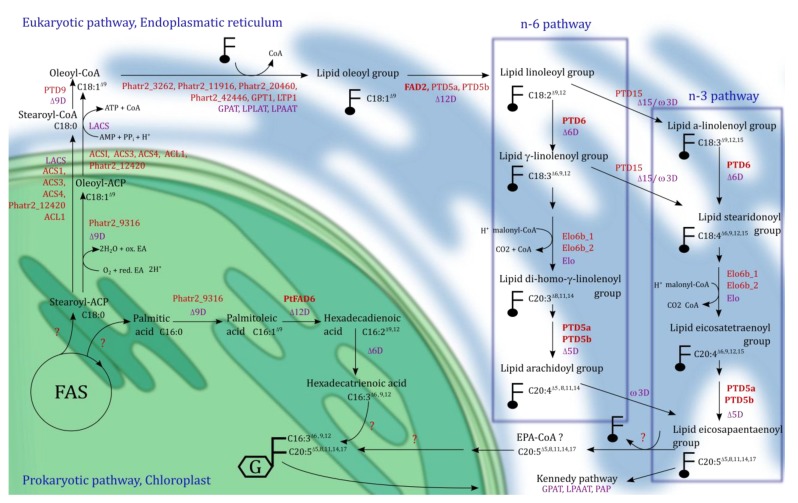
The importance of n-3 long chain polyunsaturated fatty acids (LC-PUFAs) for human health has received more focus the last decades, and the global consumption of n-3 LC-PUFA has increased. Seafood, the natural n-3 LC-PUFA source, is harvested beyond a sustainable capacity, and it is therefore imperative to develop alternative n-3 LC-PUFA sources for both eicosapentaenoic acid (EPA, 20:5n-3) and docosahexaenoic acid (DHA, 22:6n-3). Genera of algae such as Nannochloropsis, Schizochytrium, Isochrysis and Phaedactylum within the kingdom Chromista have received attention due to their ability to produce n-3 LC-PUFAs. Knowledge of LC-PUFA synthesis and its regulation in algae at the molecular level is fragmentary and represents a bottleneck for attempts to enhance the n-3 LC-PUFA levels for industrial production. In the present review, Phaeodactylum tricornutum has been used to exemplify the synthesis and compartmentalization of n-3 LC-PUFAs. Based on recent transcriptome data a co-expression network of 106 genes involved in lipid metabolism has been created. Together with recent molecular biological and metabolic studies, a model pathway for n-3 LC-PUFA synthesis in P. tricornutum has been proposed, and is compared to industrialized species of Chromista. Limitations of the n-3 LC-PUFA synthesis by enzymes such as thioesterases, elongases, acyl-CoA synthetases and acyltransferases are discussed and metabolic bottlenecks are hypothesized such as the supply of the acetyl-CoA and NADPH. A future industrialization will depend on optimization of chemical compositions and increased biomass production, which can be achieved by exploitation of the physiological potential, by selective breeding and by genetic engineering.
Figures
Similar articles
-
Towards the Industrial Production of Omega-3 Long Chain Polyunsaturated Fatty Acids from a Genetically Modified Diatom Phaeodactylum tricornutum.PLoS One. 2015 Dec 14;10(12):e0144054. doi: 10.1371/journal.pone.0144054. eCollection 2015. PLoS One. 2015. PMID: 26658738 Free PMC article.
-
Identification of two novel microalgal enzymes involved in the conversion of the omega3-fatty acid, eicosapentaenoic acid, into docosahexaenoic acid.Biochem J. 2004 Dec 1;384(Pt 2):357-66. doi: 10.1042/BJ20040970. Biochem J. 2004. PMID: 15307817 Free PMC article.
-
Production of polyunsaturated fatty acids by Schizochytrium (Aurantiochytrium) spp.Biotechnol Adv. 2022 Mar-Apr;55:107897. doi: 10.1016/j.biotechadv.2021.107897. Epub 2021 Dec 30. Biotechnol Adv. 2022. PMID: 34974158 Review.
-
Biochemical characterization of acyl-CoA:diacylglycerol acyltransferase2 from the diatom Phaeodactylum tricornutum and its potential effect on LC-PUFAs biosynthesis in planta.BMC Plant Biol. 2024 Apr 23;24(1):309. doi: 10.1186/s12870-024-05014-7. BMC Plant Biol. 2024. PMID: 38649801 Free PMC article.
-
Modifying the lipid content and composition of plant seeds: engineering the production of LC-PUFA.Appl Microbiol Biotechnol. 2015 Jan;99(1):143-54. doi: 10.1007/s00253-014-6217-2. Epub 2014 Nov 25. Appl Microbiol Biotechnol. 2015. PMID: 25417743 Free PMC article. Review.
Cited by
-
Betaine Lipid Is Crucial for Adapting to Low Temperature and Phosphate Deficiency in Nannochloropsis.Plant Physiol. 2018 May;177(1):181-193. doi: 10.1104/pp.17.01573. Epub 2018 Mar 19. Plant Physiol. 2018. PMID: 29555786 Free PMC article.
-
Fatty Acid Profiles and Production in Marine Phytoplankton.Mar Drugs. 2019 Mar 4;17(3):151. doi: 10.3390/md17030151. Mar Drugs. 2019. PMID: 30836652 Free PMC article. Review.
-
Suboptimal Temperature Acclimation Affects Kennedy Pathway Gene Expression, Lipidome and Metabolite Profile of Nannochloropsis salina during PUFA Enriched TAG Synthesis.Mar Drugs. 2018 Nov 1;16(11):425. doi: 10.3390/md16110425. Mar Drugs. 2018. PMID: 30388843 Free PMC article.
-
New Insights into Xanthophylls and Lipidomic Profile Changes Induced by Glucose Supplementation in the Marine Diatom Nitzschia laevis.Mar Drugs. 2022 Jul 14;20(7):456. doi: 10.3390/md20070456. Mar Drugs. 2022. PMID: 35877749 Free PMC article.
-
Minireview: algal natural compounds and extracts as antifoulants.J Appl Phycol. 2018;30(3):1859-1874. doi: 10.1007/s10811-017-1322-0. Epub 2017 Nov 6. J Appl Phycol. 2018. PMID: 29899600 Free PMC article. Review.
References
-
- Kelly P.B., Reiser R., Hood D.W. The origin of the marine polyunsaturated fatty acids. Composition of some marine plankton. J. Am. Oil Chem. Soc. 1959;36:104–106. doi: 10.1007/BF02639979. - DOI
-
- Yaguchi T., Tanaka S., Yokochi T., Nakahara T., Higashihara T. Production of high yields of docosahexaenoic acid by Schizochytrium sp. strain SR21. J. Am. Oil Chem. Soc. 1997;74:1431–1434. doi: 10.1007/s11746-997-0249-z. - DOI
-
- Sukenik A. Ecophysiological considerations in the optimization of eicosapentaenoic acid production by Nannochloropsis sp. (Eustigmatophyceae) Bioresour. Technol. 1991;35:263–269. doi: 10.1016/0960-8524(91)90123-2. - DOI
-
- Wan C., Bai F.-W., Zhao X.-Q. Effects of nitrogen concentration and media replacement on cell growth and lipid production of oleaginous marine microalga Nannochloropsis oceanica DUT01. Biochem. Eng. J. 2013;78:32–38. doi: 10.1016/j.bej.2013.04.014. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
