

Differential virulence and pathogenesis of West Nile viruses
- PMID: 24284878
- PMCID: PMC3856419
- DOI: 10.3390/v5112856
Differential virulence and pathogenesis of West Nile viruses
Abstract
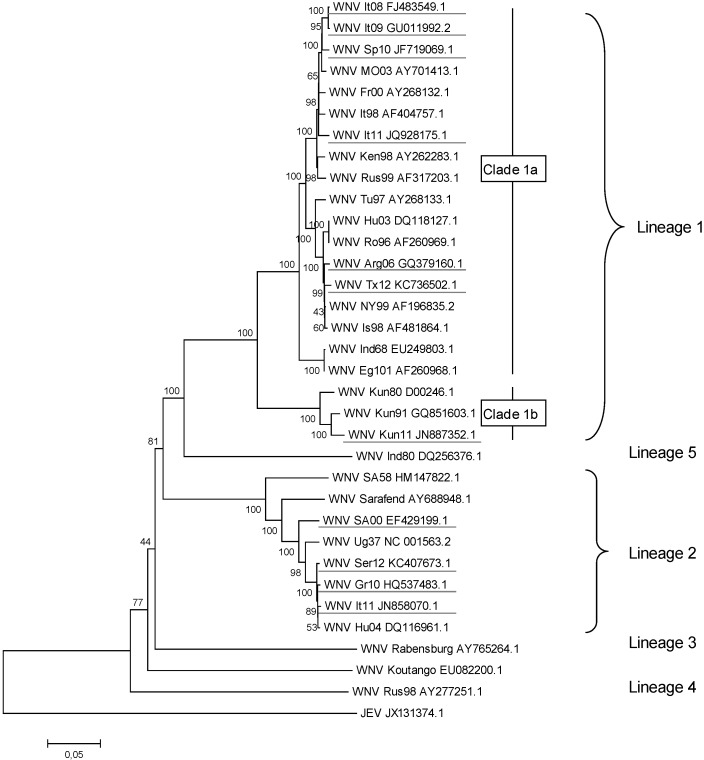
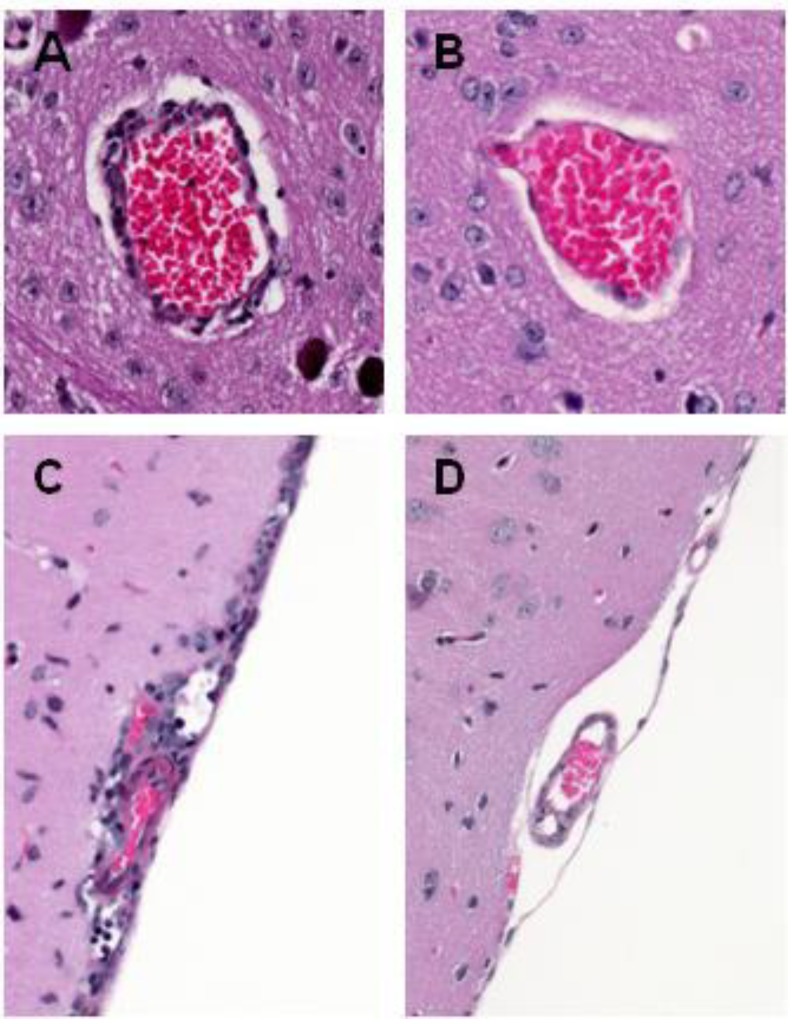
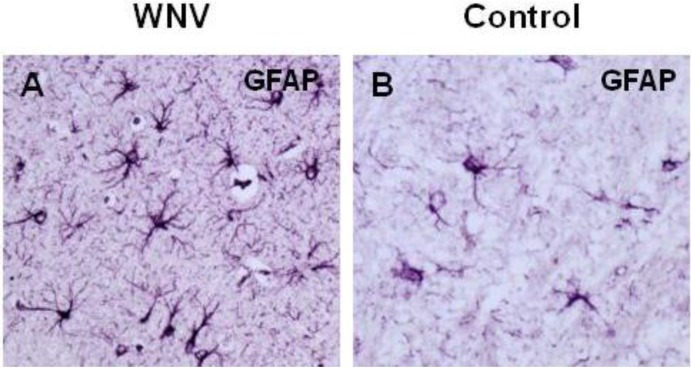
West Nile virus (WNV) is a neurotropic flavivirus that cycles between mosquitoes and birds but that can also infect humans, horses, and other vertebrate animals. In most humans, WNV infection remains subclinical. However, 20%-40% of those infected may develop WNV disease, with symptoms ranging from fever to meningoencephalitis. A large variety of WNV strains have been described worldwide. Based on their genetic differences, they have been classified into eight lineages; the pathogenic strains belong to lineages 1 and 2. Ten years ago, Beasley et al. (2002) found that dramatic differences exist in the virulence and neuroinvasion properties of lineage 1 and lineage 2 WNV strains. Further insights on how WNV interacts with its hosts have recently been gained; the virus acts either at the periphery or on the central nervous system (CNS), and these observed differences could help explain the differential virulence and neurovirulence of WNV strains. This review aims to summarize the current state of knowledge on factors that trigger WNV dissemination and CNS invasion as well as on the inflammatory response and CNS damage induced by WNV. Moreover, we will discuss how WNV strains differentially interact with the innate immune system and CNS cells, thus influencing WNV pathogenesis.
Figures

References
-
- Smithburn K.C., Hughes T.P., Burke A.W., Paul J.H. A neurotropic virus isolated from the blood of a native of Uganda. Am. J. Trop. Med. Hyg. 1940;20:471–492.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
