

Terminator disparity contributes to stereo matching for eye movements and perception
- PMID: 24285893
- PMCID: PMC3841453
- DOI: 10.1523/JNEUROSCI.3332-13.2013
Terminator disparity contributes to stereo matching for eye movements and perception
Abstract
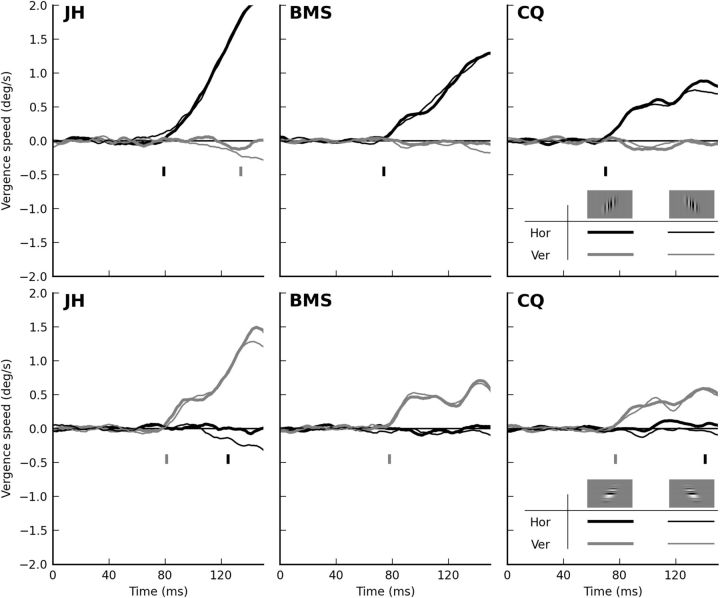
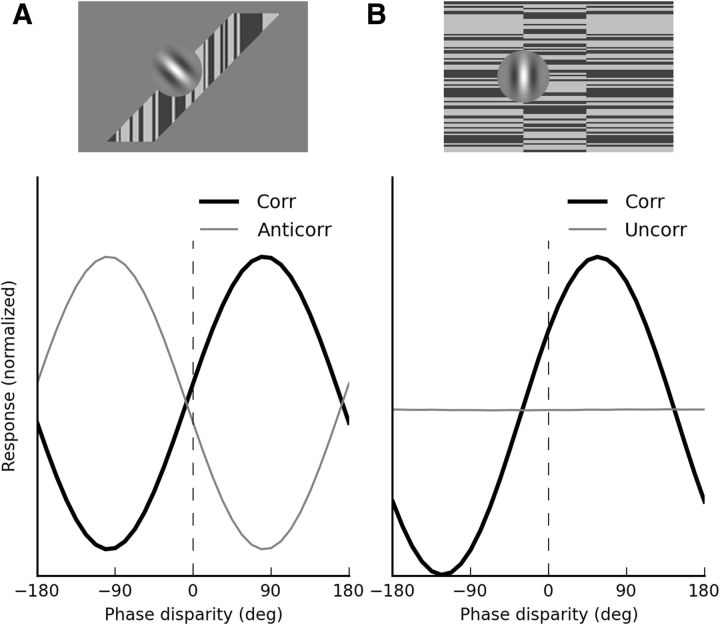
In the context of motion detection, the endings (or terminators) of 1-D features can be detected as 2-D features, affecting the perceived direction of motion of the 1-D features (the barber-pole illusion) and the direction of tracking eye movements. In the realm of binocular disparity processing, an equivalent role for the disparity of terminators has not been established. Here we explore the stereo analogy of the barber-pole stimulus, applying disparity to a 1-D noise stimulus seen through an elongated, zero-disparity, aperture. We found that, in human subjects, these stimuli induce robust short-latency reflexive vergence eye movements, initially in the direction orthogonal to the 1-D features, but shortly thereafter in the direction predicted by the disparity of the terminators. In addition, these same stimuli induce vivid depth percepts, which can only be attributed to the disparity of line terminators. When the 1-D noise patterns are given opposite contrast in the two eyes (anticorrelation), both components of the vergence response reverse sign. Finally, terminators drive vergence even when the aperture is defined by a texture (as opposed to a contrast) boundary. These findings prove that terminators contribute to stereo matching, and constrain the type of neuronal mechanisms that might be responsible for the detection of terminator disparity.
Figures





Similar articles
-
Vergence eye movements in response to binocular disparity without depth perception.Nature. 1997 Sep 18;389(6648):283-6. doi: 10.1038/38496. Nature. 1997. PMID: 9305842
-
Responses of primary visual cortical neurons to binocular disparity without depth perception.Nature. 1997 Sep 18;389(6648):280-3. doi: 10.1038/38487. Nature. 1997. PMID: 9305841
-
Neuronal Representation of 3-D Space in the Primary Visual Cortex and Control of Eye Movements.Perception. 2015;44(8-9):995-1006. doi: 10.1177/0301006615594930. Epub 2015 Aug 14. Perception. 2015. PMID: 26562914 Review.
-
Single-unit activity in cortical area MST associated with disparity-vergence eye movements: evidence for population coding.J Neurophysiol. 2001 May;85(5):2245-66. doi: 10.1152/jn.2001.85.5.2245. J Neurophysiol. 2001. PMID: 11353039
-
Mechanisms of stereopsis in monkey visual cortex.Cereb Cortex. 1995 May-Jun;5(3):193-204. doi: 10.1093/cercor/5.3.193. Cereb Cortex. 1995. PMID: 7613075 Review.
Cited by
-
Binocular Summation for Reflexive Eye Movements: A Potential Diagnostic Tool for Stereodeficiencies.Invest Ophthalmol Vis Sci. 2018 Dec 3;59(15):5816-5822. doi: 10.1167/iovs.18-24520. Invest Ophthalmol Vis Sci. 2018. PMID: 30521669 Free PMC article.
-
Binocular summation for reflexive eye movements.J Vis. 2018 Apr 1;18(4):7. doi: 10.1167/18.4.7. J Vis. 2018. PMID: 29621384 Free PMC article.
-
Suppression and Contrast Normalization in Motion Processing.J Neurosci. 2017 Nov 8;37(45):11051-11066. doi: 10.1523/JNEUROSCI.1572-17.2017. Epub 2017 Oct 10. J Neurosci. 2017. PMID: 29018158 Free PMC article.
References
-
- Anzai A, Ohzawa I, Freeman RD. Neural mechanisms for processing binocular information I. Simple cells. J Neurophysiol. 1999;82:891–908. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources