

Review
doi: 10.1186/gb4143.
Characterizing 5-methylcytosine in the mammalian epitranscriptome
- PMID: 24286375
- PMCID: PMC4053770
- DOI: 10.1186/gb4143
Item in Clipboard
Review
Characterizing 5-methylcytosine in the mammalian epitranscriptome
Genome Biol.
.
Abstract
The post-transcriptional modification 5-methylcytosine (m5C) occurs in a wide range of coding and non-coding RNAs. We describe transcriptome-wide approaches to capture the global m5C RNA methylome. We also discuss the potential functions of m5C in RNA and compare them to 6-methyladenosine modifications.
Figures
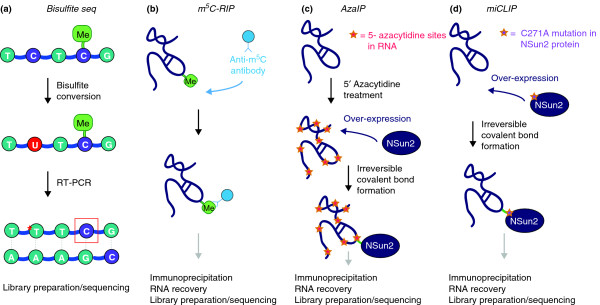
A comparison of four techniques used to identify transcriptome-wide m5C sites. (a) Bisulfite sequencing, (b) m5C-RIP, (c) Aza-IP and (d) miCLIP can all be used to detect m5C sites. The asterisk in (a) indicates bisulfite-mediated conversion of unmethylated cytosines in RNA, and the red box indicates non-conversion due to the presence of cytosine methylation. In Aza-IP, the cytidine analog 5-azacytidine sites are randomly introduced into nascent RNA following cellular exposure to 5-azacytidine, and in miCLIP, the NSun2 protein is engineered to harbor the C271A mutation. Whereas transcriptome-wide bisulfite sequencing does not enrich for methylation targets, m5C-RIP, Aza-IP and miCLIP do so via immunoprecipitation, with Aza-IP and miCLIP utilizing covalently bound protein-RNA complexes. The introduction of a covalent bond between protein and RNA in Aza-IP and miCLIP is dependent on the modified sites introduced into RNA substrates and the NSun2 protein, respectively.
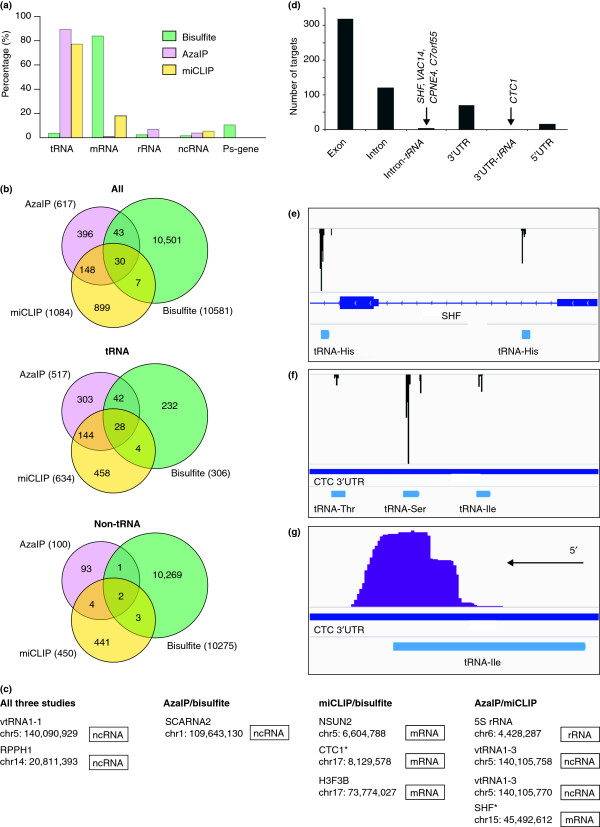
A comparison of m5C target sites identified by three transcriptome-wide approaches (Additional file1) [[18,23,24]]. The bisulfite study identifies the global pattern of m5C methylation in the genome, while the AzaIP and miCLIP data shown focus specifically on Nsun2-mediated methylation. (a) Types of transcripts found to be methylated by each technique. (b) Comparison of the genomic locations of individual methylation sites found by each of the three approaches. The Venn diagrams show the overlap between all sites (upper panel), the sites that map to tRNAs (middle panel), and non-tRNA sites (lower panel). (c) Non-tRNA target sites found in at least two of the three studies, which represent high-confidence sites. The genomic location (hg19) of methylated cytosines is given. * = target sites that overlap with tRNA locations. (d) The proportion of miCLIP-identified targets occurring in exons/introns/UTRs of mRNAs, including those sites overlapping with tRNA genes. (e) Genome browser view of the miCLIP-identified SHF mRNA methylation. Predicted methylation sites are indicated by the black peaks. Note that methylation may occur either in an SHF mRNA intron or within a tRNA, which is transcribed from the same genomic DNA strand. (f) miCLIP data showing identified sites within the CTC1 3′ UTR. The CTC1 3′ UTR contains three tRNA genes predicted to be transcribed from the same genomic DNA strand. The miCLIP predicted methylation sites (black peaks) overlapped specifically with these three regions. (g) An example showing that all miCLIP sequence reads, indicated by purple bars, only extend slightly beyond the annotated tRNA genes. It is most likely that these represent extensions only into the 3′ trailer sequences of tRNAs, and that tRNA rather than mRNA methylation is being detected.
References
-
- Machnicka MA, Milanowska K, Osman Oglou O, Purta E, Kurkowska M, Olchowik A, Januszewski W, Kalinowski S, Dunin-Horkawicz S, Rother KM, Helm M, Bujnicki JM, Grosjean H. MODOMICS: a database of RNA modification pathways - 2013 update. Nucleic Acids Res. 2013;14:D262–D267. doi: 10.1093/nar/gks1007. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
