

Differential regulation of the apoptotic machinery during megakaryocyte differentiation and platelet production by inhibitor of apoptosis protein Livin
- PMID: 24287698
- PMCID: PMC3847330
- DOI: 10.1038/cddis.2013.454
Differential regulation of the apoptotic machinery during megakaryocyte differentiation and platelet production by inhibitor of apoptosis protein Livin
Abstract
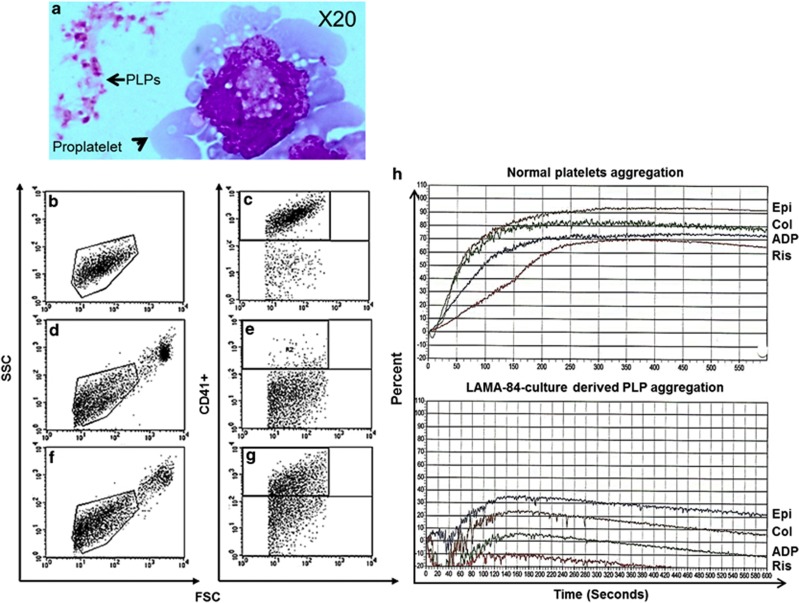
Livin is a member of the inhibitor of apoptosis proteins (IAP) family of intracellular antiapoptotic proteins that act by binding and inhibiting caspases. Upon strong apoptotic stimuli, it is then specifically cleaved by caspases to produce a truncated protein (tLivin) with a paradoxical proapoptotic activity. Intriguingly, we have detected robust protein levels of Livin in normal mature bone marrow megakaryocyte (MK) and platelets. To evaluate the potential role of Livin in thrombopoiesis, we used the human BCR-ABL+ cell line, LAMA-84, and cord blood CD34+ cells to induce differentiation toward MKs. Upon differentiation, induced by phorbol myristate acetate and concurrent with increase in Livin protein expression, LAMA-84 cells formed functional platelet-like particles. Livin overexpression in CD34+ progenitor cells induced higher endoreplication in the MKs generated. Furthermore, overexpression of Livin increased the ability of both primary MKs and differentiated LAMA-84 cells to produce functional platelets. In the differentiated LAMA-84 cells, we observed accumulation of proapoptotic tLivin concomitant with increased caspase-3 activity. Downregulation of Livin with small interfering RNA in both leukemic and primary MK cells decreased their ability to produce functional platelets. We suggest that Livin has a role in thrombopoiesis by regulating the apoptotic and antiapoptotic balance in MK endoreplication and platelet production.
Figures






References
-
- Ravid K, Lu J, Zimmet JM, Jones MR. Roads to polyploidy: The megakaryocyte example. J Cell Physiol. 2002;190:7–20. - PubMed
-
- Deutsch VR, Tomer A. Advances in megakaryocytopoiesis and thrombopoiesis: from bench to bedside. Br J Haematol. 2013;161:778–793. - PubMed
-
- Handagama PJ, Feldman BF, Jain NC, Farver TB, Kono CS. In vitro platelet release by rat megakaryocytes: effect of metabolic inhibitors and cytoskeletal disrupting agents. Am J Vet Res. 1987;48:1142–1146. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
