

Hexanucleotide repeats in ALS/FTD form length-dependent RNA foci, sequester RNA binding proteins, and are neurotoxic
- PMID: 24290757
- PMCID: PMC3898469
- DOI: 10.1016/j.celrep.2013.10.049
Hexanucleotide repeats in ALS/FTD form length-dependent RNA foci, sequester RNA binding proteins, and are neurotoxic
Abstract
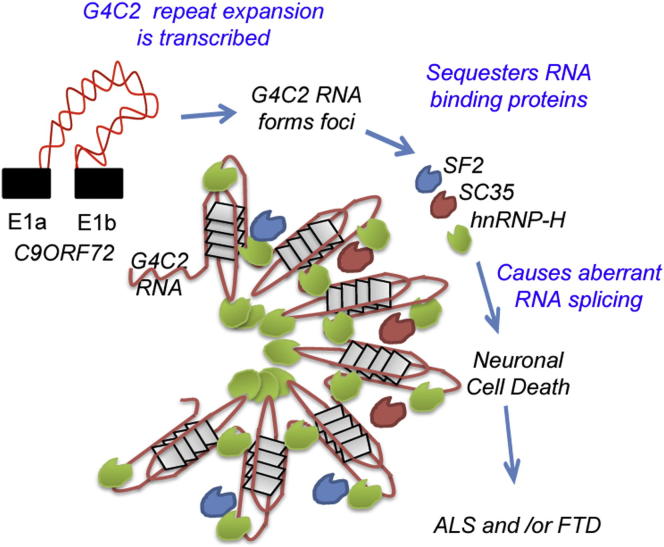
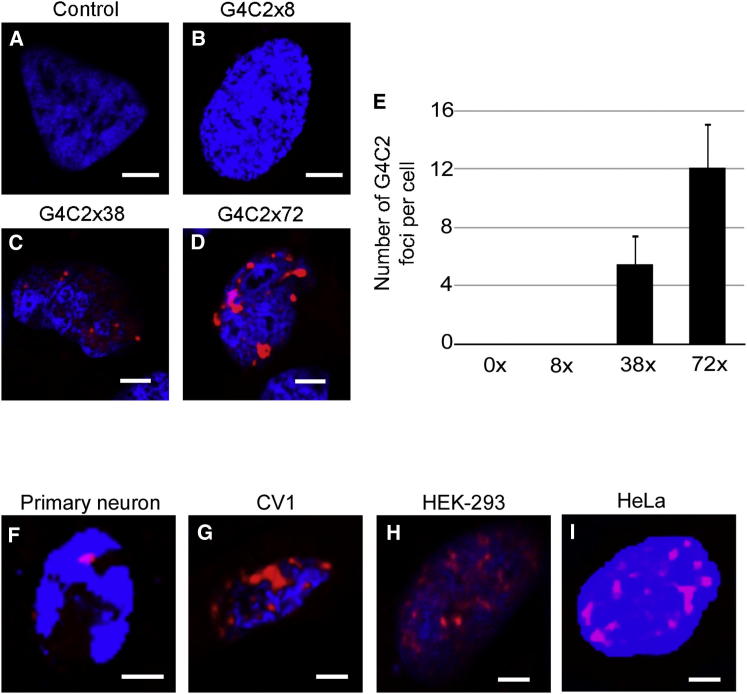
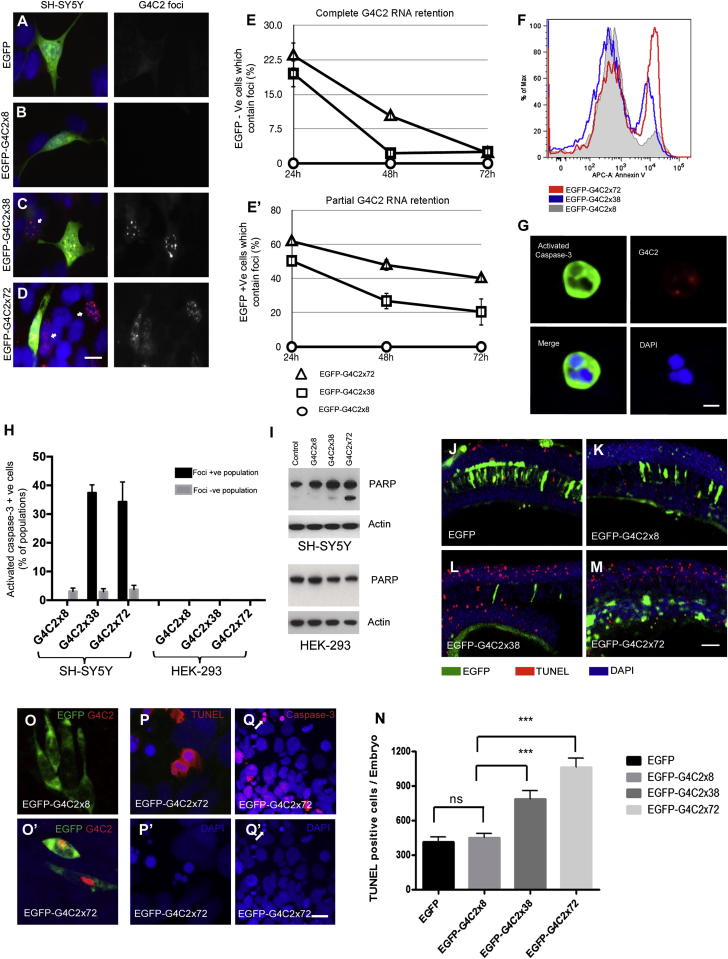
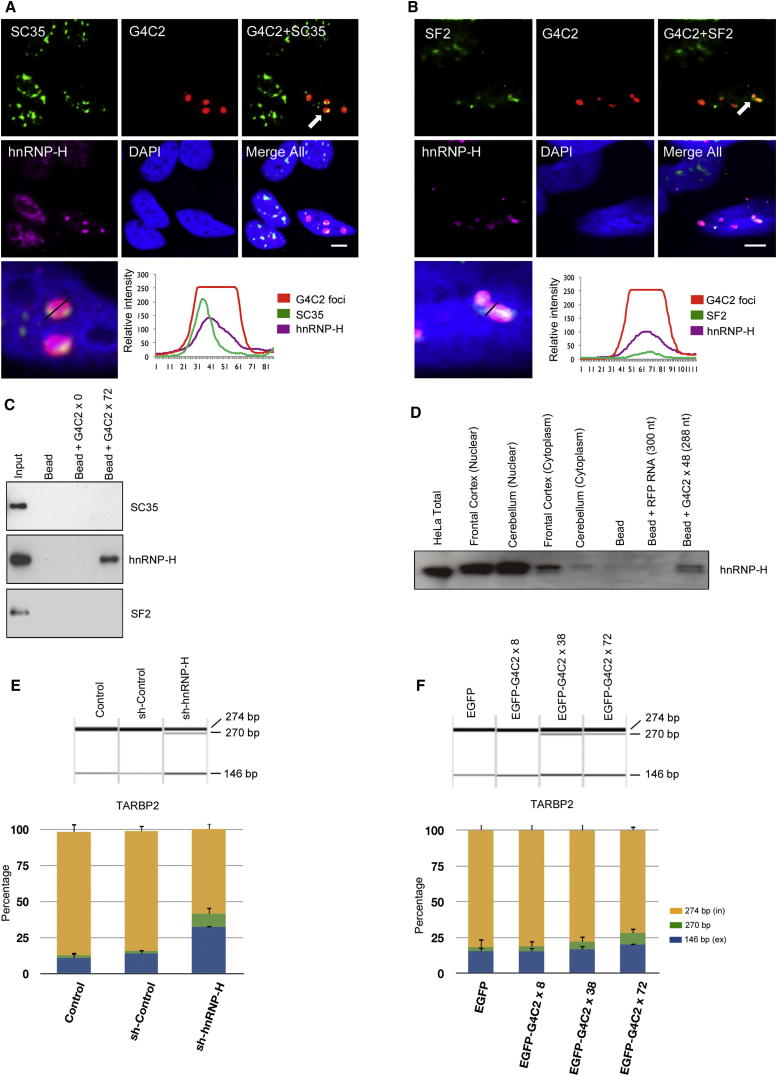
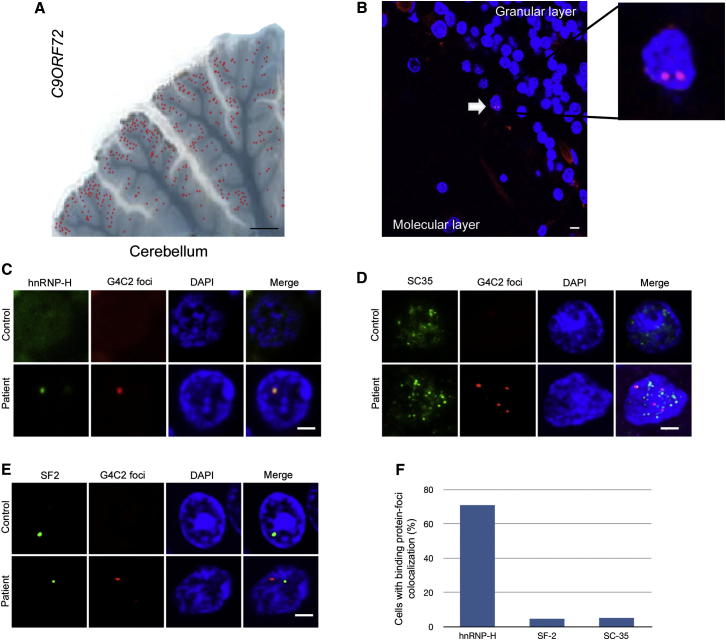
The GGGGCC (G4C2) intronic repeat expansion within C9ORF72 is the most common genetic cause of amyotrophic lateral sclerosis (ALS) and frontotemporal dementia (FTD). Intranuclear neuronal RNA foci have been observed in ALS and FTD tissues, suggesting that G4C2 RNA may be toxic. Here, we demonstrate that the expression of 38× and 72× G4C2 repeats form intranuclear RNA foci that initiate apoptotic cell death in neuronal cell lines and zebrafish embryos. The foci colocalize with a subset of RNA binding proteins, including SF2, SC35, and hnRNP-H in transfected cells. Only hnRNP-H binds directly to G4C2 repeats following RNA immunoprecipitation, and only hnRNP-H colocalizes with 70% of G4C2 RNA foci detected in C9ORF72 mutant ALS and FTD brain tissues. We show that expanded G4C2 repeats are potently neurotoxic and bind hnRNP-H and other RNA binding proteins. We propose that RNA toxicity and protein sequestration may disrupt RNA processing and contribute to neurodegeneration.
Copyright © 2013 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Al-Sarraj S., King A., Troakes C., Smith B., Maekawa S., Bodi I., Rogelj B., Al-Chalabi A., Hortobágyi T., Shaw C.E. p62 positive, TDP-43 negative, neuronal cytoplasmic and intranuclear inclusions in the cerebellum and hippocampus define the pathology of C9orf72-linked FTLD and MND/ALS. Acta Neuropathol. 2011;122:691–702. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
