

Should evolutionary geneticists worry about higher-order epistasis?
- PMID: 24290990
- PMCID: PMC4313208
- DOI: 10.1016/j.gde.2013.10.007
Should evolutionary geneticists worry about higher-order epistasis?
Abstract
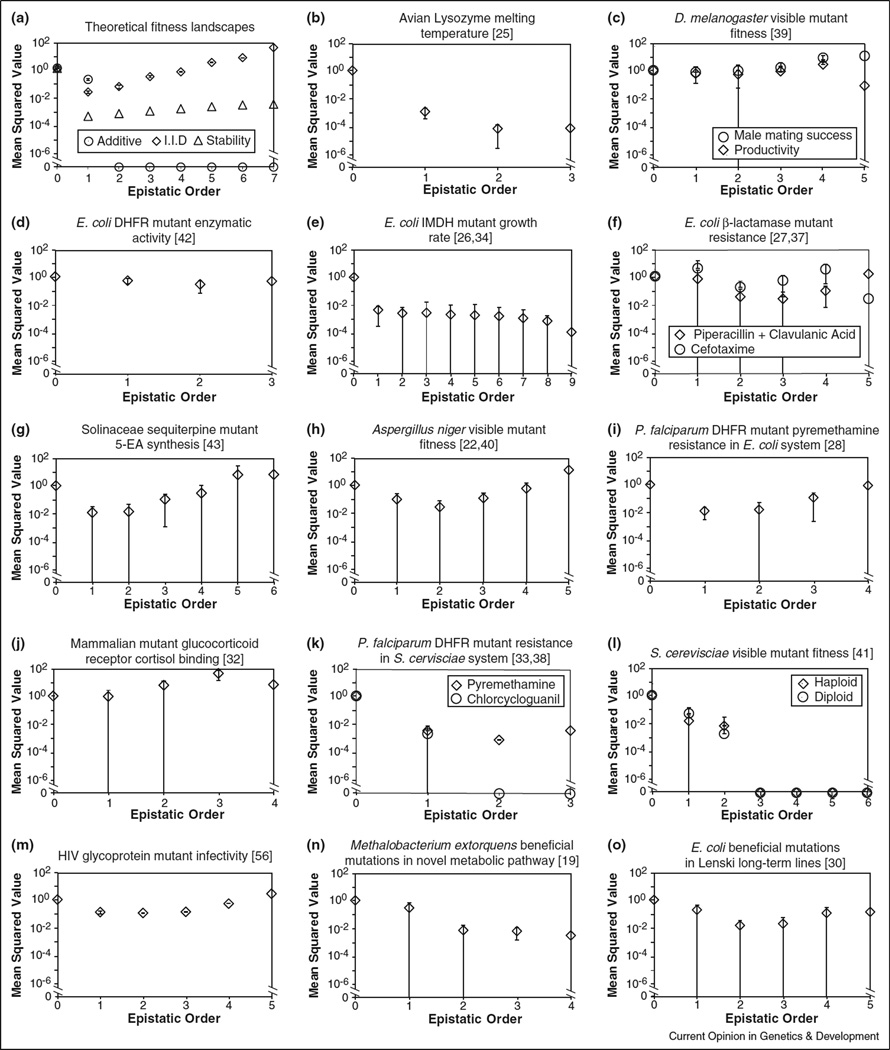
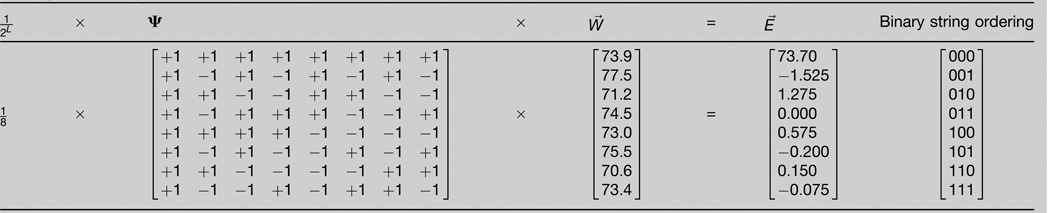
Natural selection drives evolving populations up the fitness landscape, the projection from nucleotide sequence space to organismal reproductive success. While it has long been appreciated that topographic complexities on fitness landscapes can arise only as a consequence of epistatic interactions between mutations, evolutionary genetics has mainly focused on epistasis between pairs of mutations. Here we propose a generalization to the classical population genetic treatment of pairwise epistasis that yields expressions for epistasis among arbitrary subsets of mutations of all orders (pairwise, three-way, etc.). Our approach reveals substantial higher-order epistasis in almost every published fitness landscape. Furthermore we demonstrate that higher-order epistasis is critically important in two systems we know best. We conclude that higher-order epistasis deserves empirical and theoretical attention from evolutionary geneticists.
Copyright © 2013 Elsevier Ltd. All rights reserved.
Figures
References
-
- Bateson W. Mendel’s Principles of Heredity. Cambridge: Cambridge University Press; 1909.
-
-
Pettersson M, Besnier F, Siegal PB, Carlborg O. Replication and exploration of high-order epistasis using a large advanced intercross line pedigree. PLoS Genet. 2011;7:e1002180. Genetic architecture of growth in chicken was found to exhibit third-order epistasis in very large pedigree experimental design
-
-
-
Wang Y, Arenas CD, Stoebel DM, Cooper TF. Genetic background affects epistatic interactions between two beneficial mutations. Biol Lett. 2013;9:20120328. Beneficial mutations in E. coli first isolated from Richard Lenski’s long-term experimental evolution lines were placed into two natural isolates and found to exhibit distinctly different patterns of epistasis
-
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
