

Role of myelin plasticity in oscillations and synchrony of neuronal activity
- PMID: 24291730
- PMCID: PMC4037390
- DOI: 10.1016/j.neuroscience.2013.11.007
Role of myelin plasticity in oscillations and synchrony of neuronal activity
Abstract
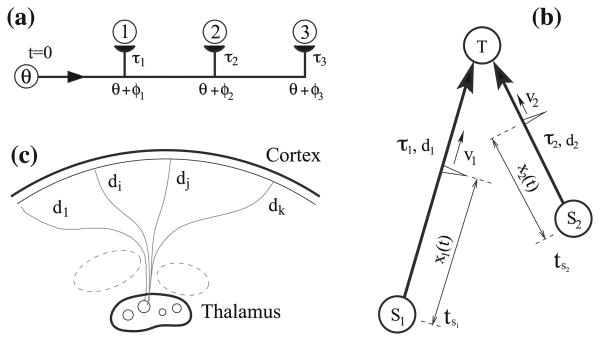
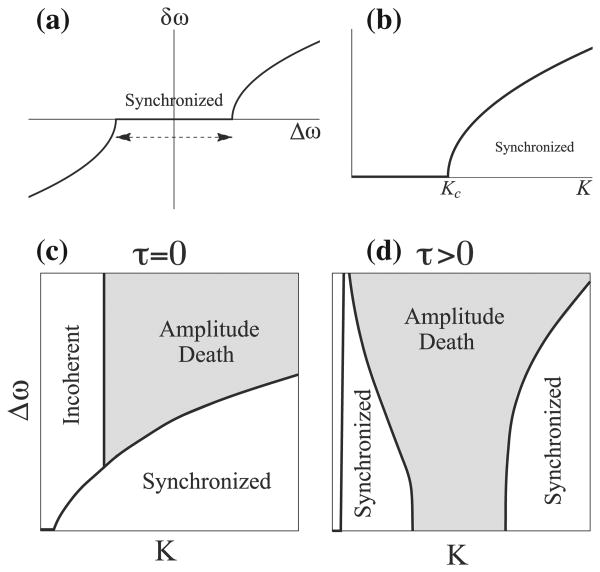
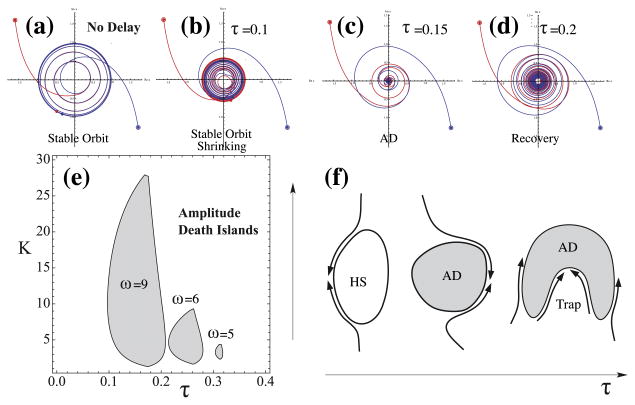
Conduction time is typically ignored in computational models of neural network function. Here we consider the effects of conduction delays on the synchrony of neuronal activity and neural oscillators, and evaluate the consequences of allowing conduction velocity (CV) to be regulated adaptively. We propose that CV variation, mediated by myelin, could provide an important mechanism of activity-dependent nervous system plasticity. Even small changes in CV, resulting from small changes in myelin thickness or nodal structure, could have profound effects on neuronal network function in terms of spike-time arrival, oscillation frequency, oscillator coupling, and propagation of brain waves. For example, a conduction delay of 5ms could change interactions of two coupled oscillators at the upper end of the gamma frequency range (∼100Hz) from constructive to destructive interference; delays smaller than 1ms could change the phase by 30°, significantly affecting signal amplitude. Myelin plasticity, as another form of activity-dependent plasticity, is relevant not only to nervous system development but also to complex information processing tasks that involve coupling and synchrony among different brain rhythms. We use coupled oscillator models with time delays to explore the importance of adaptive time delays and adaptive synaptic strengths. The impairment of activity-dependent myelination and the loss of adaptive time delays may contribute to disorders where hyper- and hypo-synchrony of neuronal firing leads to dysfunction (e.g., dyslexia, schizophrenia, epilepsy).
Keywords: activity-dependent myelination; conduction velocity and delays; coupled oscillators; oscillations; synchronization; white matter plasticity.
Published by Elsevier Ltd.
Figures
References
-
- Abraham H, Vincze A, Jewgenow I, Beszpremi B, Kravjak A, Gomori E, Seress L. Myelination in the human hippocampal formation form midgestation to adulthood. Int J Dev Neurosci. 2010;28:401–410. - PubMed
-
- Ainsworth M, Lee S, Cunningham MO, Traub RD, Kopell NJ, Whittington MA. Rates and rhythms: a synergistic view of frequency and temporal coding in neuronal networks. Neuron. 2012;75:572–583. - PubMed
-
- Aronson DG, Ermentrout GB, Kopell N. Amplitude response of coupled oscillators. Phys D Nonlinear Phenom. 1990;41:403–449.
-
- Bakiri Y, Burzomato V, Frugier G, Hamilton NB, Karadottir R, Attwell D. Glutamatergic signaling in the brain's white matter. Neuroscience. 2009;158:266–274. - PubMed
-
- Bar-Eli K. On the stability of coupled chemical oscillators. Phys D Nonlinear Phenom. 1985;14:242–252.
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
