

Thiol-based redox regulation in sexual plant reproduction: new insights and perspectives
- PMID: 24294217
- PMCID: PMC3827552
- DOI: 10.3389/fpls.2013.00465
Thiol-based redox regulation in sexual plant reproduction: new insights and perspectives
Abstract
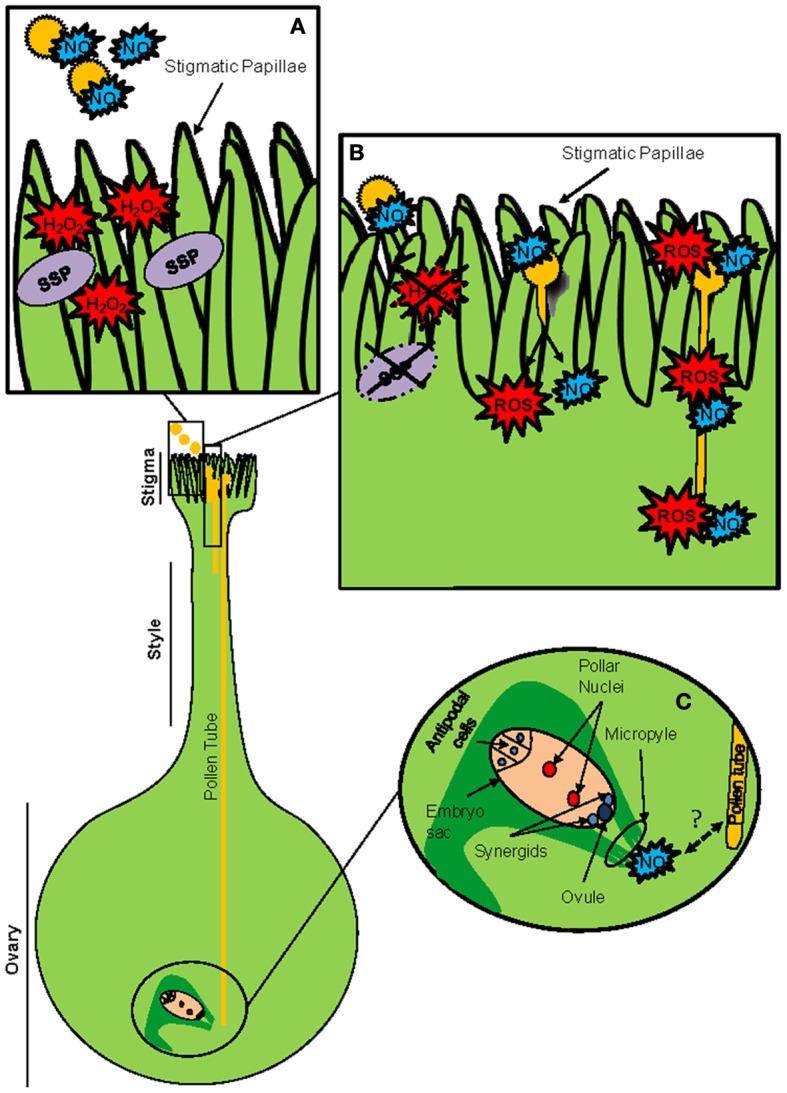
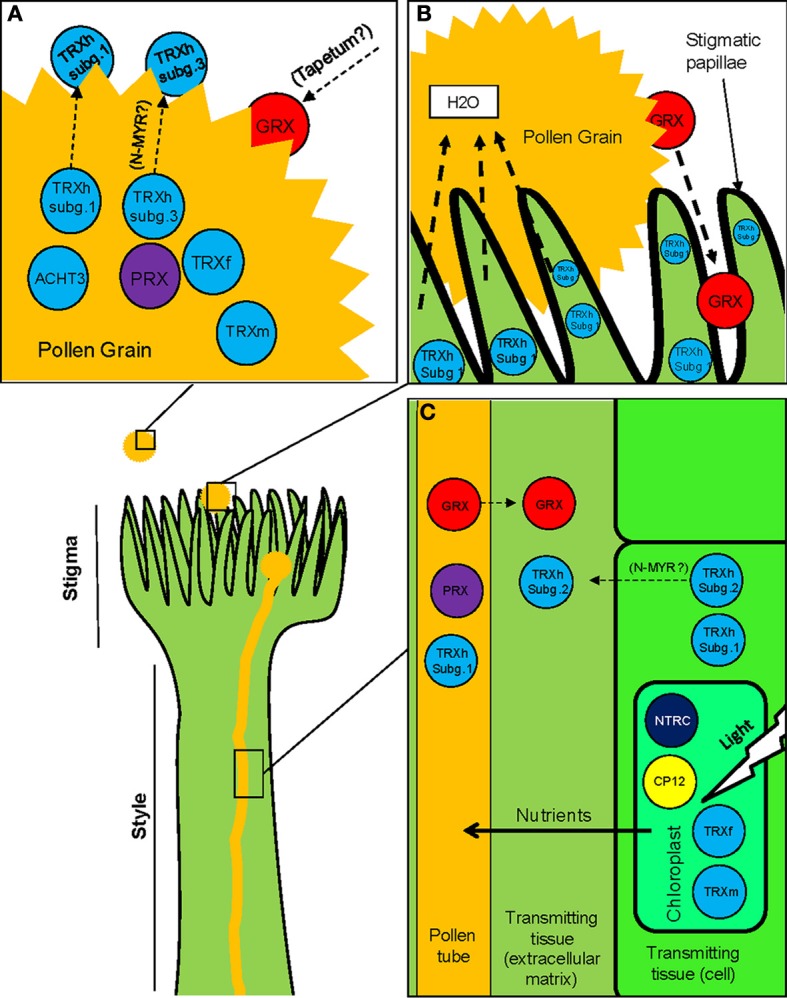
The success of sexual reproduction in plants involves (i) the proper formation of the plant gametophytes (pollen and embryo sac) containing the gametes, (ii) the accomplishment of specific interactions between pollen grains and the stigma, which subsequently lead to (iii) the fusion of the gametes and eventually to (iv) the seed setting. Owing to the lack of mobility, plants have developed specific regulatory mechanisms to control all developmental events underlying the sexual plant reproduction according to environmental challenges. Over the last decade, redox regulation and signaling have come into sight as crucial mechanisms able to manage critical stages during sexual plant reproduction. This regulation involves a complex redox network which includes reactive oxygen species (ROS), reactive nitrogen species (RNS), glutathione and other classic buffer molecules or antioxidant proteins, and some thiol/disulphide-containing proteins belonging to the thioredoxin superfamily, like glutaredoxins (GRXs) or thioredoxins (TRXs). These proteins participate as critical elements not only in the switch between the mitotic to the meiotic cycle but also at further developmental stages of microsporogenesis. They are also implicated in the regulation of pollen rejection as the result of self-incompatibility. In addition, they display precise space-temporal patterns of expression and are present in specific localizations like the stigmatic papillae or the mature pollen, although their functions and subcellular localizations are not clear yet. In this review we summarize insights and perspectives about the presence of thiol/disulphide-containing proteins in plant reproduction, taking into account the general context of the cell redox network.
Keywords: glutaredoxin; pistil; pollen; pollen-pistil interaction; redox regulation; sexual plant reproduction; stigma; thioredoxin.
Figures

Similar articles
-
Involvement of thiol-based mechanisms in plant development.Biochim Biophys Acta. 2015 Aug;1850(8):1479-96. doi: 10.1016/j.bbagen.2015.01.023. Epub 2015 Feb 9. Biochim Biophys Acta. 2015. PMID: 25676896 Review.
-
Gametophyte interaction and sexual reproduction: how plants make a zygote.Int J Dev Biol. 2005;49(5-6):615-32. doi: 10.1387/ijdb.052023lb. Int J Dev Biol. 2005. PMID: 16096969 Review.
-
NADPH Oxidase-Dependent Superoxide Production in Plant Reproductive Tissues.Front Plant Sci. 2016 Mar 31;7:359. doi: 10.3389/fpls.2016.00359. eCollection 2016. Front Plant Sci. 2016. PMID: 27066025 Free PMC article. Review.
-
The Molecular and Cellular Regulation of Brassicaceae Self-Incompatibility and Self-Pollen Rejection.Int Rev Cell Mol Biol. 2019;343:1-35. doi: 10.1016/bs.ircmb.2018.05.011. Epub 2018 Jun 22. Int Rev Cell Mol Biol. 2019. PMID: 30712670 Review.
-
Cellular localization of ROS and NO in olive reproductive tissues during flower development.BMC Plant Biol. 2010 Feb 24;10:36. doi: 10.1186/1471-2229-10-36. BMC Plant Biol. 2010. PMID: 20181244 Free PMC article.
Cited by
-
Arabidopsis ABCG28 is required for the apical accumulation of reactive oxygen species in growing pollen tubes.Proc Natl Acad Sci U S A. 2019 Jun 18;116(25):12540-12549. doi: 10.1073/pnas.1902010116. Epub 2019 May 31. Proc Natl Acad Sci U S A. 2019. PMID: 31152136 Free PMC article.
-
A disulfide bond A-like oxidoreductase is a strong candidate gene for self-incompatibility in apricot (Prunus armeniaca) pollen.J Exp Bot. 2017 Nov 2;68(18):5069-5078. doi: 10.1093/jxb/erx336. J Exp Bot. 2017. PMID: 29036710 Free PMC article.
-
Proteomic profiling reveals insights into Triticeae stigma development and function.J Exp Bot. 2014 Nov;65(20):6069-80. doi: 10.1093/jxb/eru350. Epub 2014 Aug 28. J Exp Bot. 2014. PMID: 25170101 Free PMC article.
-
Sexual reproduction in land plants: an evolutionary perspective.Plant Reprod. 2025 May 12;38(2):12. doi: 10.1007/s00497-025-00522-4. Plant Reprod. 2025. PMID: 40355640 Free PMC article. Review.
-
Effects of Regulated Deficit Irrigation on Amino Acid Profiles and Their Derived Volatile Compounds in Cabernet Sauvignon (Vitis vinifera L.) Grapes and Wines.Molecules. 2018 Aug 9;23(8):1983. doi: 10.3390/molecules23081983. Molecules. 2018. PMID: 30096898 Free PMC article.
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources