

Deciphering the role of aspartate and prephenate aminotransferase activities in plastid nitrogen metabolism
- PMID: 24296073
- PMCID: PMC3875828
- DOI: 10.1104/pp.113.232462
Deciphering the role of aspartate and prephenate aminotransferase activities in plastid nitrogen metabolism
Abstract
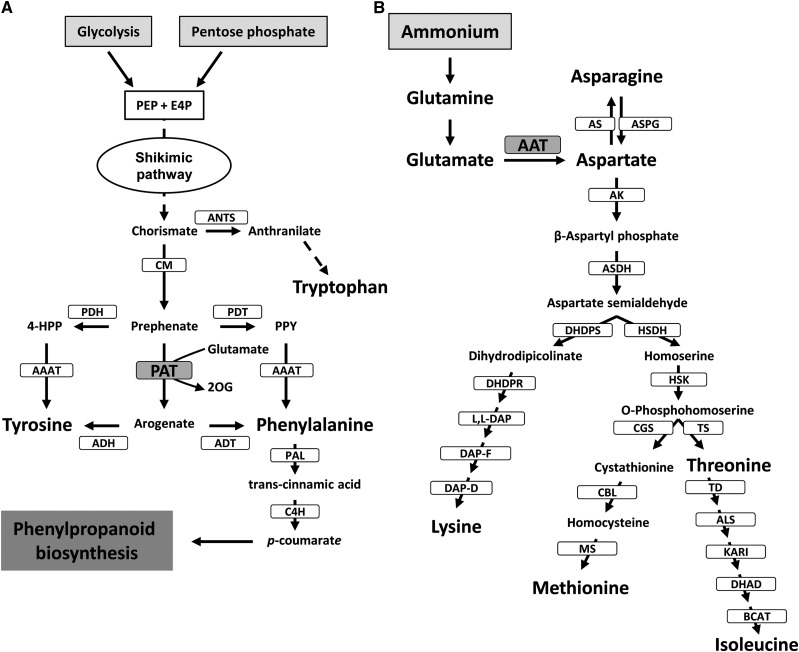
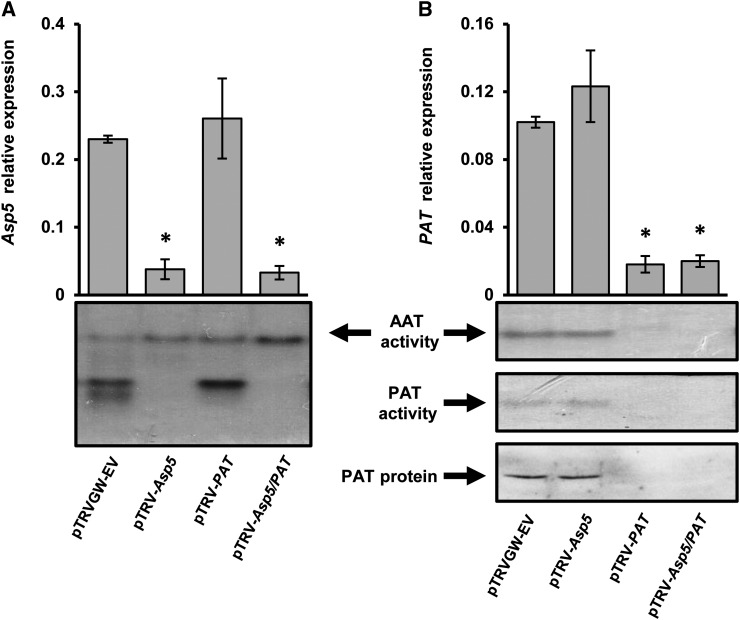
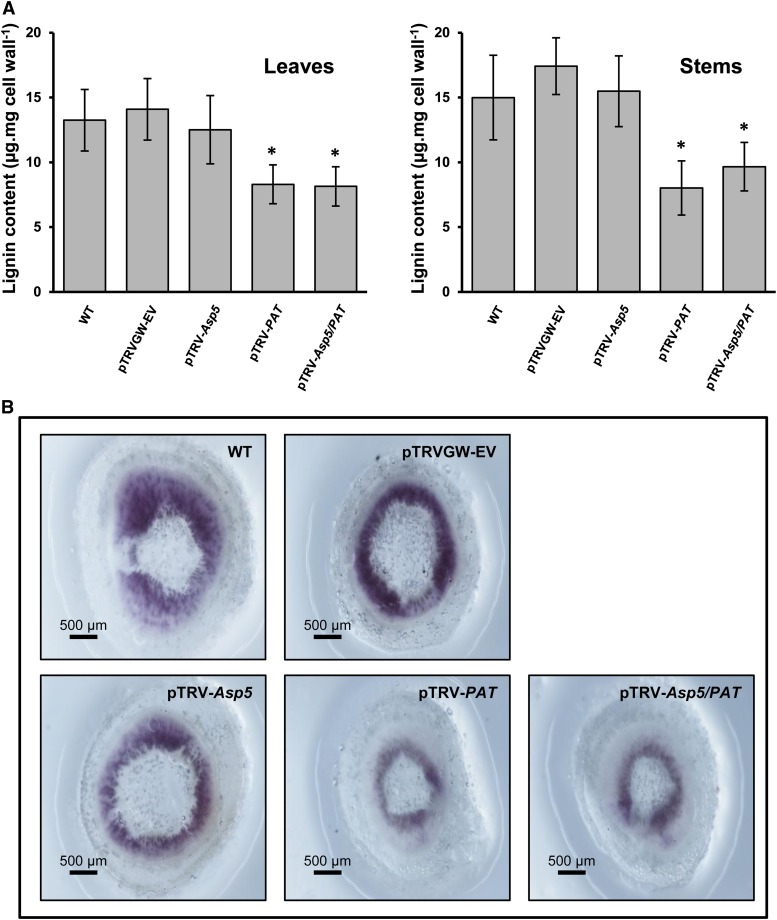
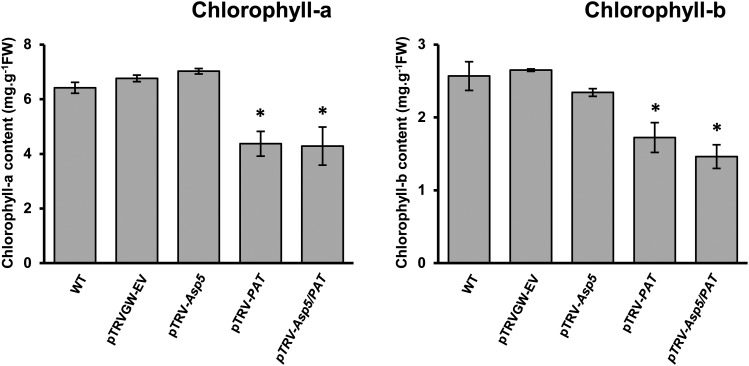
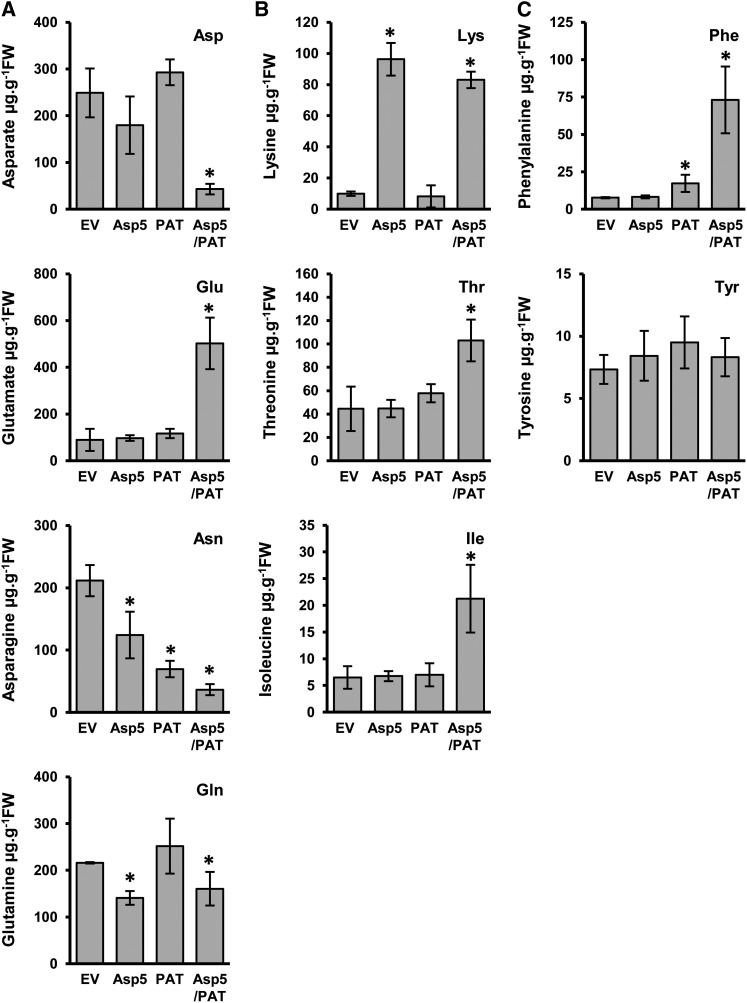
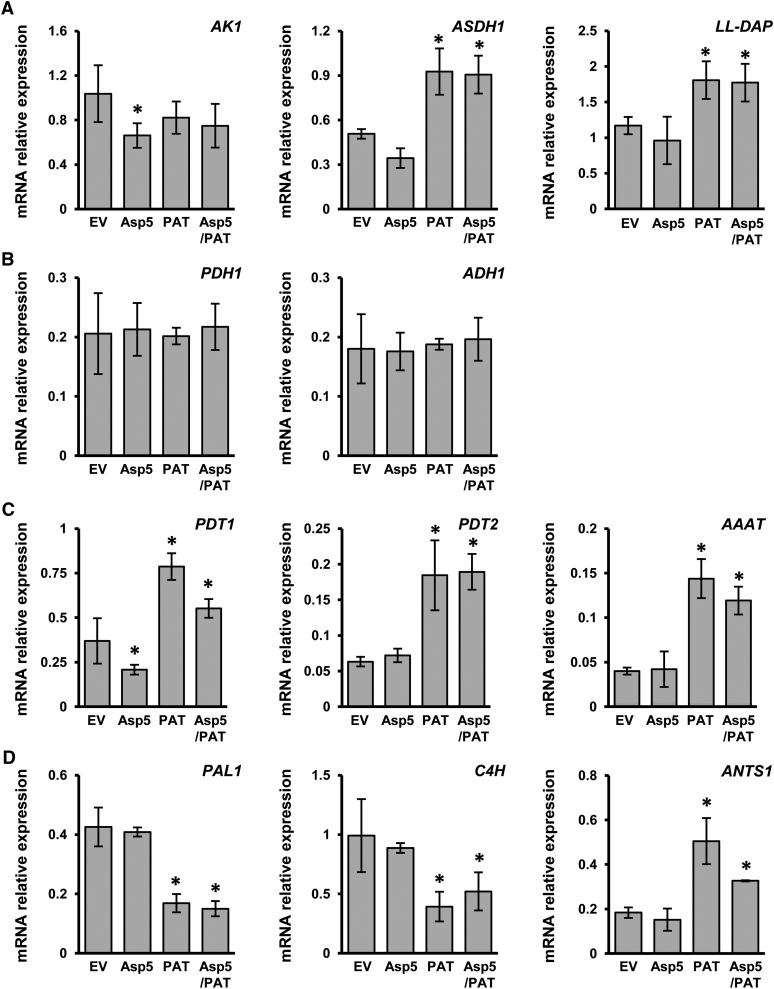
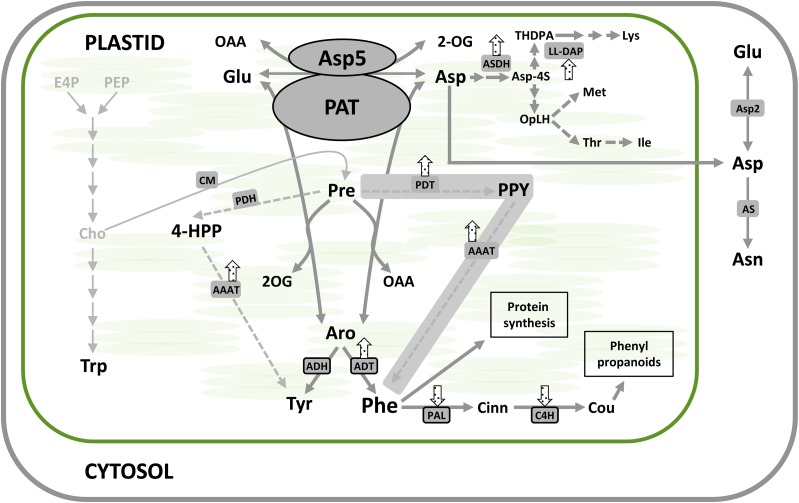
Chloroplasts and plastids of nonphotosynthetic plant cells contain two aspartate (Asp) aminotransferases: a eukaryotic type (Asp5) and a prokaryotic-type bifunctional enzyme displaying Asp and prephenate aminotransferase activities (PAT). We have identified the entire Asp aminotransferase gene family in Nicotiana benthamiana and isolated and cloned the genes encoding the isoenzymes with plastidic localization: NbAsp5 and NbPAT. Using a virus-induced gene silencing approach, we obtained N. benthamiana plants silenced for NbAsp5 and/or NbPAT. Phenotypic and metabolic analyses were conducted in silenced plants to investigate the specific roles of these enzymes in the biosynthesis of essential amino acids within the plastid. The NbAsp5 silenced plants had no changes in phenotype, exhibiting similar levels of free Asp and glutamate as control plants, but contained diminished levels of asparagine and much higher levels of lysine. In contrast, the suppression of NbPAT led to a severe reduction in growth and strong chlorosis symptoms. NbPAT silenced plants exhibited extremely reduced levels of asparagine and were greatly affected in their phenylalanine metabolism and lignin deposition. Furthermore, NbPAT suppression triggered a transcriptional reprogramming in plastid nitrogen metabolism. Taken together, our results indicate that NbPAT has an overlapping role with NbAsp5 in the biosynthesis of Asp and a key role in the production of phenylalanine for the biosynthesis of phenylpropanoids. The analysis of NbAsp5/NbPAT cosilenced plants highlights the central role of both plastidic aminotransferases in nitrogen metabolism; however, only NbPAT is essential for plant growth and development.
Figures

References
-
- Azevedo RA, Lancien M, Lea PJ. (2006) The aspartic acid metabolic pathway, an exciting and essential pathway in plants. Amino Acids 30: 143–162 - PubMed
-
- Brauc S, De Vooght E, Claeys M, Höfte M, Angenon G. (2011) Influence of over-expression of cytosolic aspartate aminotransferase on amino acid metabolism and defence responses against Botrytis cinerea infection in Arabidopsis thaliana. J Plant Physiol 168: 1813–1819 - PubMed
-
- Buchanan BB, Gruissem W, Jones RL (2000) Biochemistry and Molecular Biology of Plants. American Society of Plant Physiologists, Rockville, MD
MeSH terms
Substances
LinkOut - more resources
Other Literature Sources