

The roles of evolutionarily conserved functional modules in cilia-related trafficking
- PMID: 24296415
- PMCID: PMC4016715
- DOI: 10.1038/ncb2888
The roles of evolutionarily conserved functional modules in cilia-related trafficking
Abstract
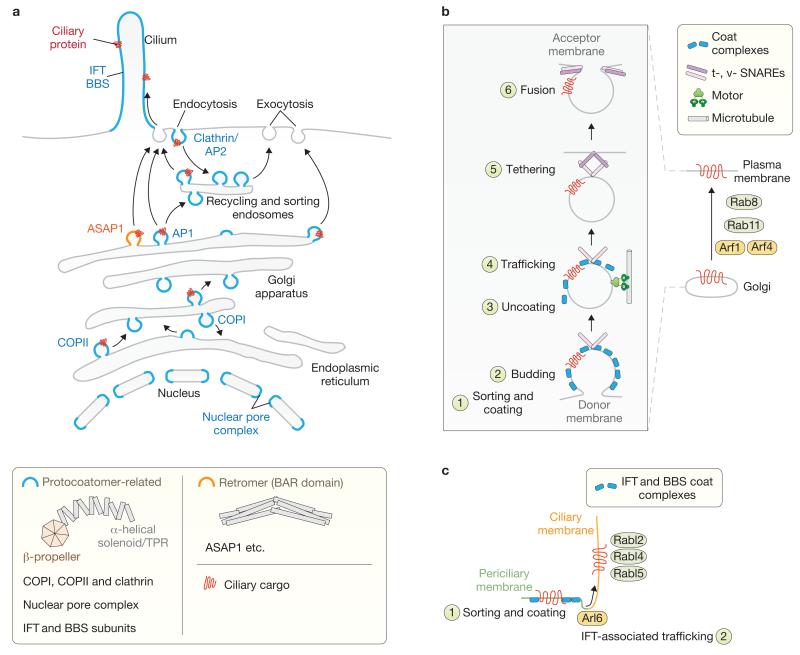
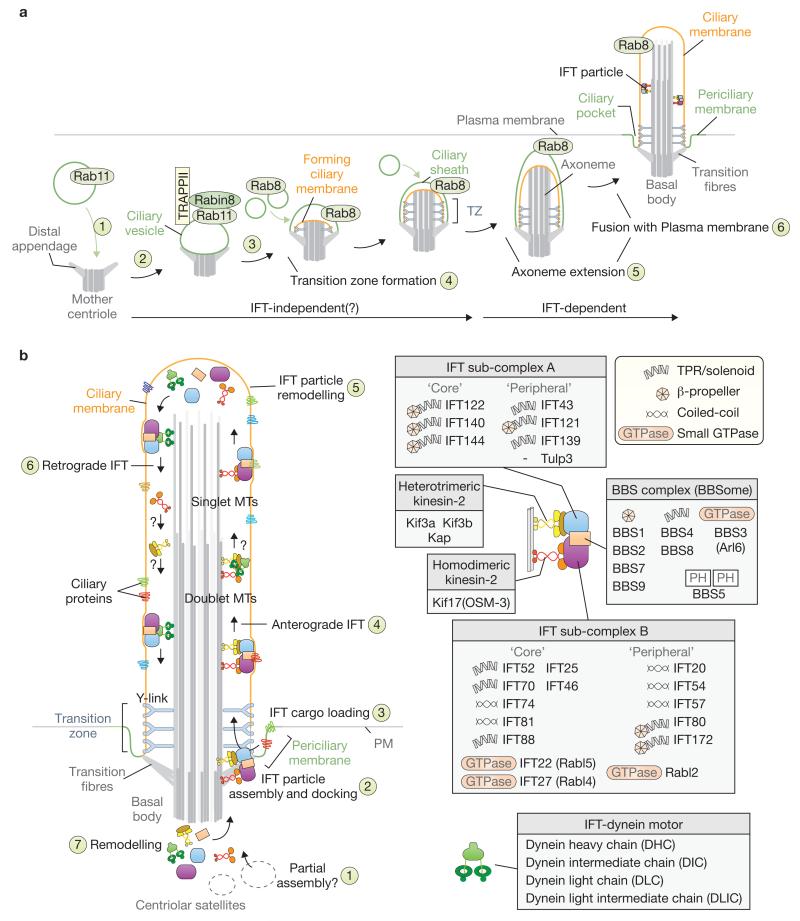
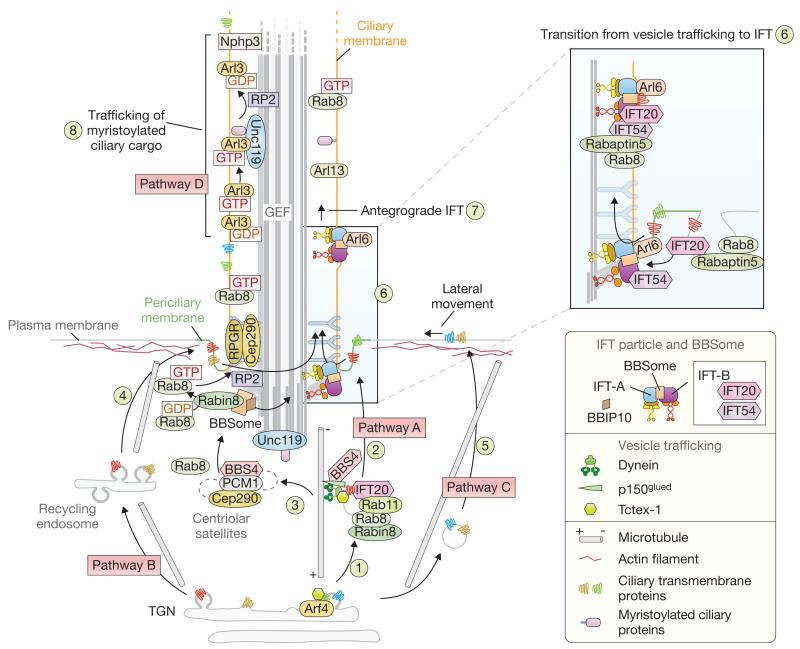
Cilia are present across most eukaryotic phyla and have diverse sensory and motility roles in animal physiology, cell signalling and development. Their biogenesis and maintenance depend on vesicular and intraciliary (intraflagellar) trafficking pathways that share conserved structural and functional modules. The functional units of the interconnected pathways, which include proteins involved in membrane coating as well as small GTPases and their accessory factors, were first experimentally associated with canonical vesicular trafficking. These components are, however, ancient, having been co-opted by the ancestral eukaryote to establish the ciliary organelle, and their study can inform us about ciliary biology in higher organisms.
Figures
References
-
- Cavalier-Smith T. Predation and eukaryote cell origins: a coevolutionary perspective. Int. J. Biochem. Cell Biol. 2009;41:307–322. - PubMed
-
- Bornens M. The centrosome in cells and organisms. Science. 2012;335:422–426. - PubMed
-
- Fisch C, Dupuis-Williams P. Ultrastructure of cilia and flagella – back to the future! Biol. Cell. 2011;103:249–270. - PubMed
-
- Ishikawa H, Marshall WF. Ciliogenesis: building the cell’s antenna. Nat. Rev. Mol. Cell Biol. 2011;12:222–234. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
