

Epistasis and quantitative traits: using model organisms to study gene-gene interactions
- PMID: 24296533
- PMCID: PMC3918431
- DOI: 10.1038/nrg3627
Epistasis and quantitative traits: using model organisms to study gene-gene interactions
Abstract
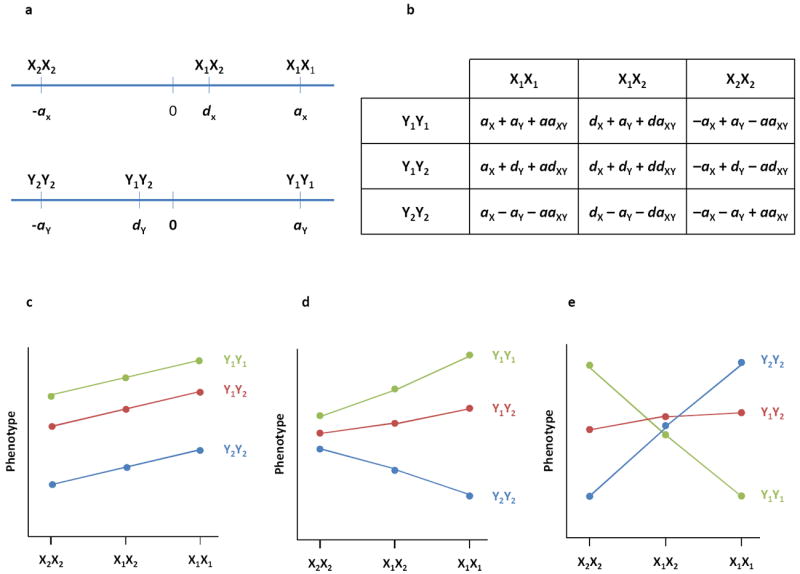
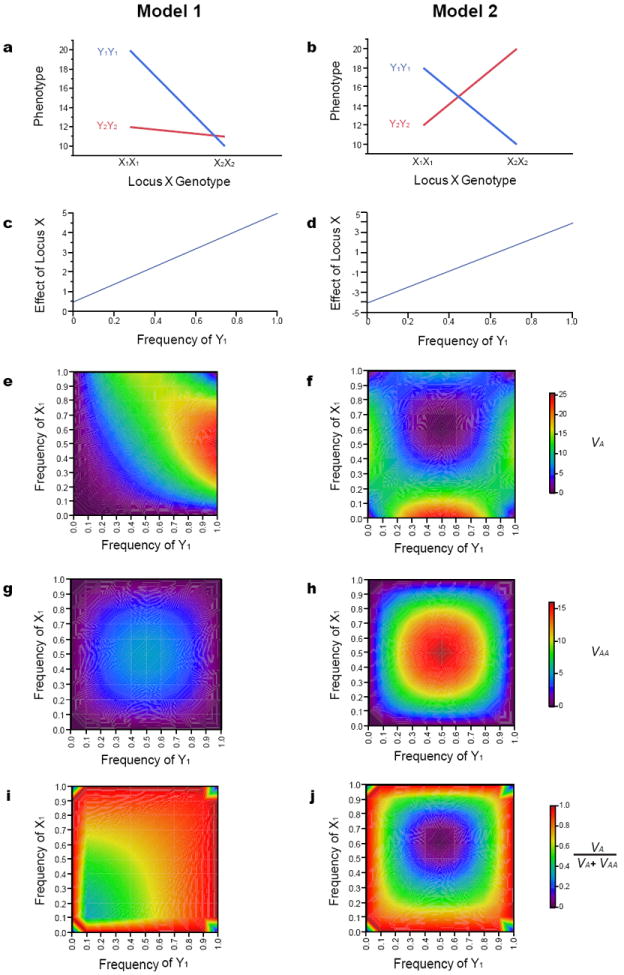
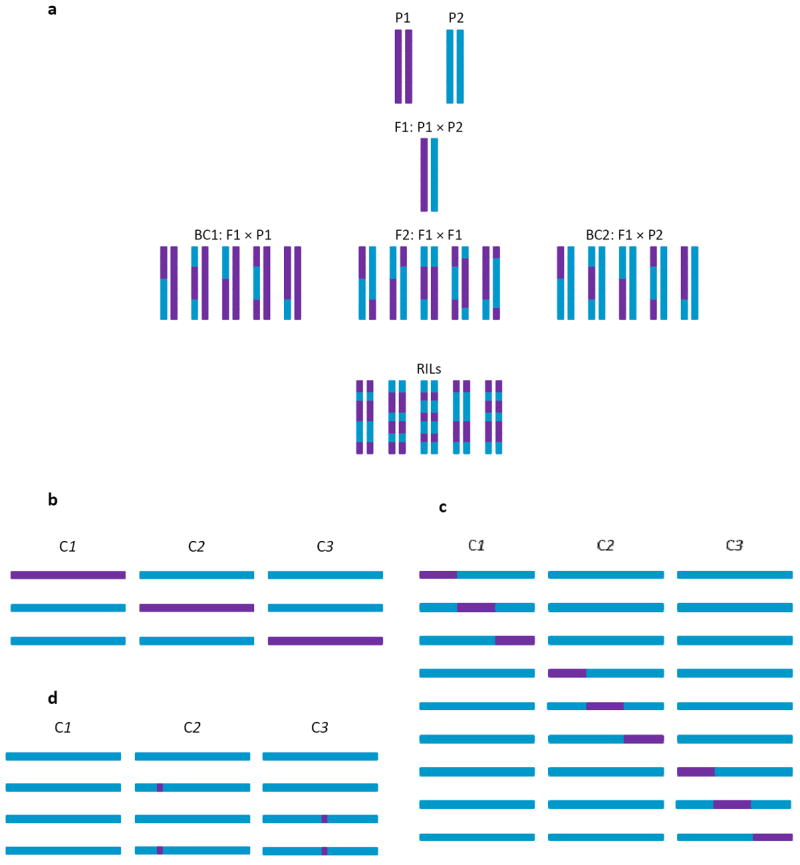
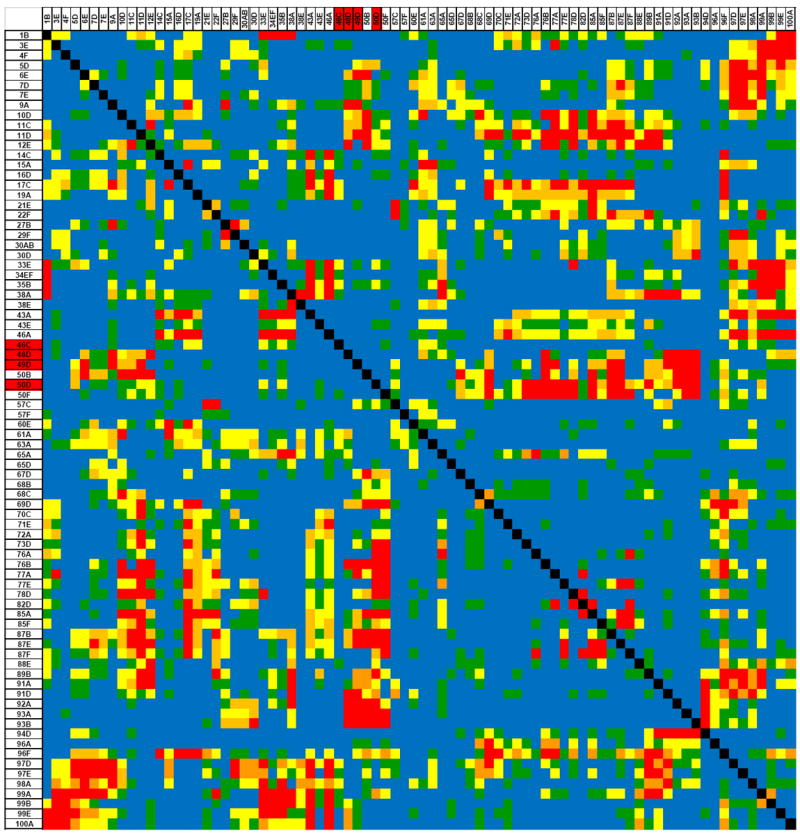
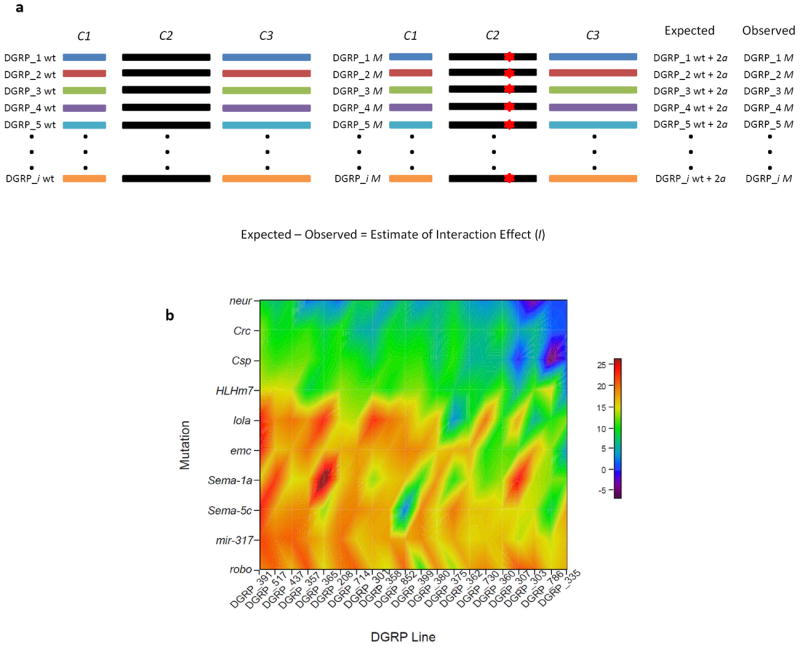
The role of epistasis in the genetic architecture of quantitative traits is controversial, despite the biological plausibility that nonlinear molecular interactions underpin the genotype-phenotype map. This controversy arises because most genetic variation for quantitative traits is additive. However, additive variance is consistent with pervasive epistasis. In this Review, I discuss experimental designs to detect the contribution of epistasis to quantitative trait phenotypes in model organisms. These studies indicate that epistasis is common, and that additivity can be an emergent property of underlying genetic interaction networks. Epistasis causes hidden quantitative genetic variation in natural populations and could be responsible for the small additive effects, missing heritability and the lack of replication that are typically observed for human complex traits.
Figures
References
-
- Kauffman SA. The Origins of Order. Oxford University Press; New York: 1993.
-
- Phillips PC. Epistasis - the essential role of gene interactions in the structure and evolution of genetic systems. Nat Rev Genet. 2008;9:855–867. A comprehensive review describing the importance of studying gene interactions with respect to dissecting regulatory pathways, mapping the genetic basis of complex traits and understanding the structure and evolution of complex systems. - PMC - PubMed
-
- Waddington CH. Canalization of development and the inheritance of acquired characters. Nature. 1942;150:563–565. - PubMed
-
- Waddington CH. The Strategy of Genes. George Allen and Unwin; London: 1957.
-
- Dobzhansky T. Genetics and the Origin of Species. Columbia University Press; New York: 1937.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
