

Hypocretin (orexin) regulates glutamate input to fast-spiking interneurons in layer V of the Fr2 region of the murine prefrontal cortex
- PMID: 24297328
- PMCID: PMC4397574
- DOI: 10.1093/cercor/bht326
Hypocretin (orexin) regulates glutamate input to fast-spiking interneurons in layer V of the Fr2 region of the murine prefrontal cortex
Abstract
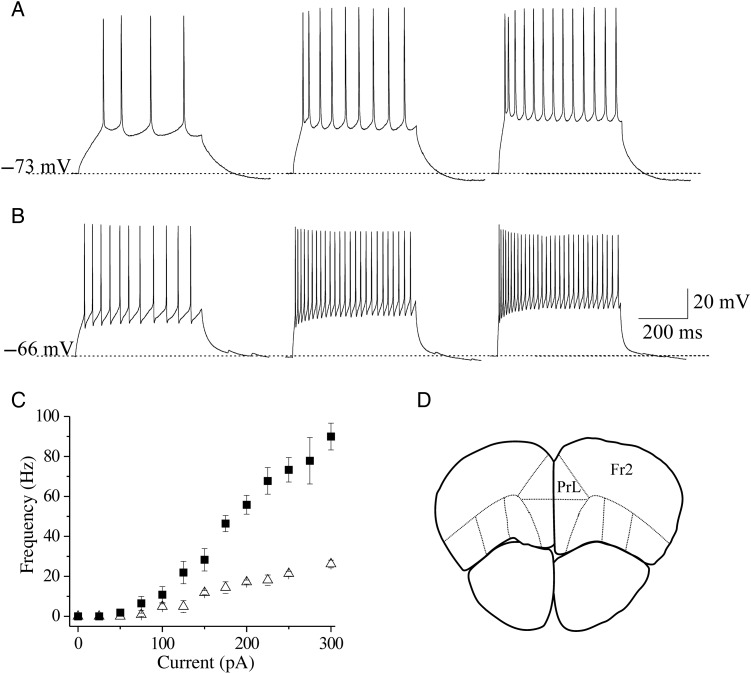
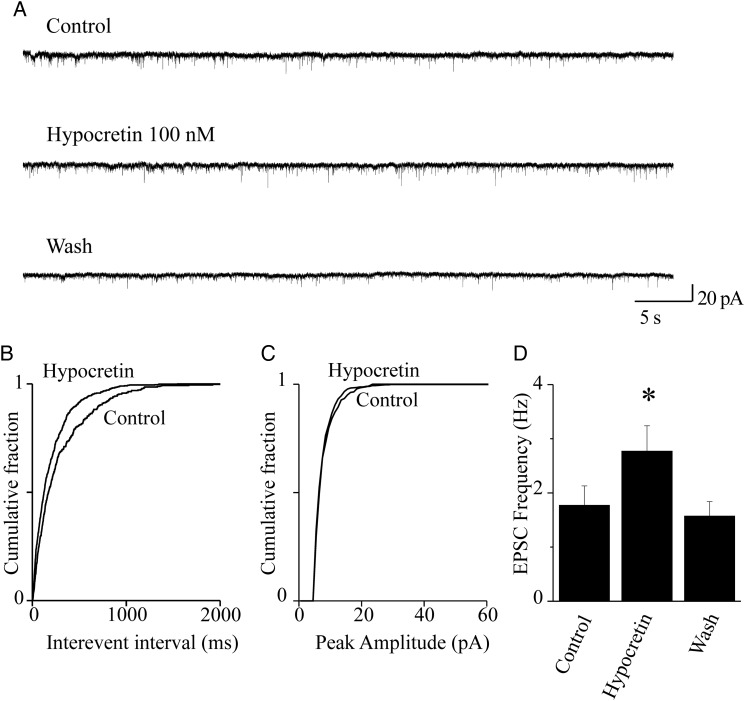
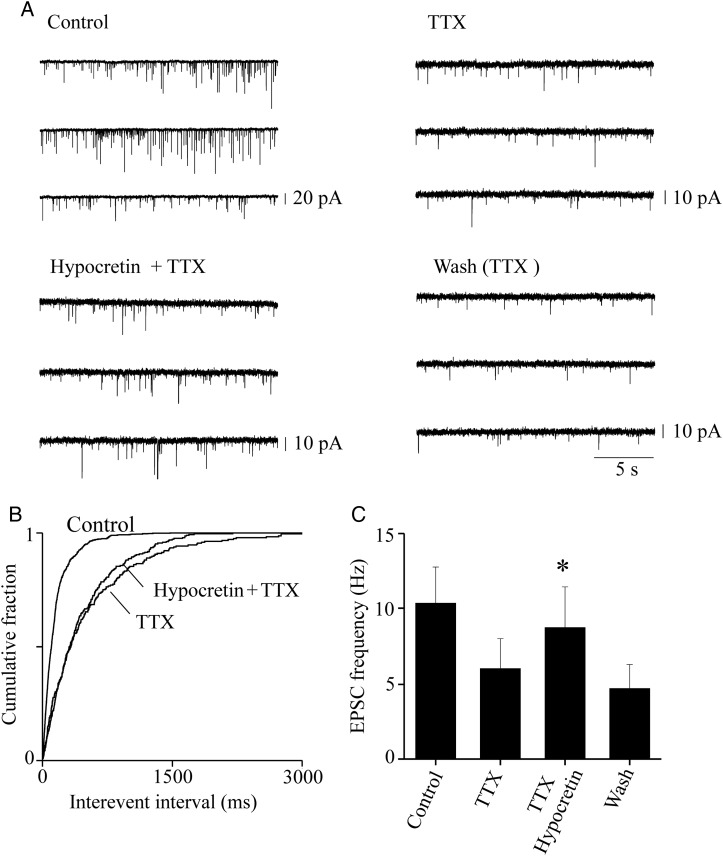
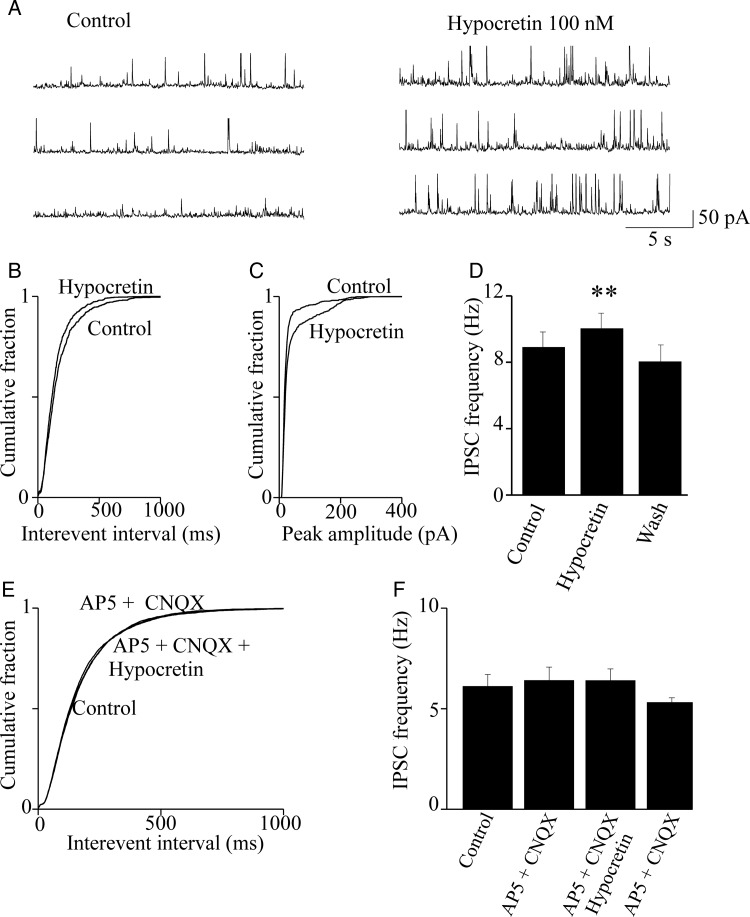
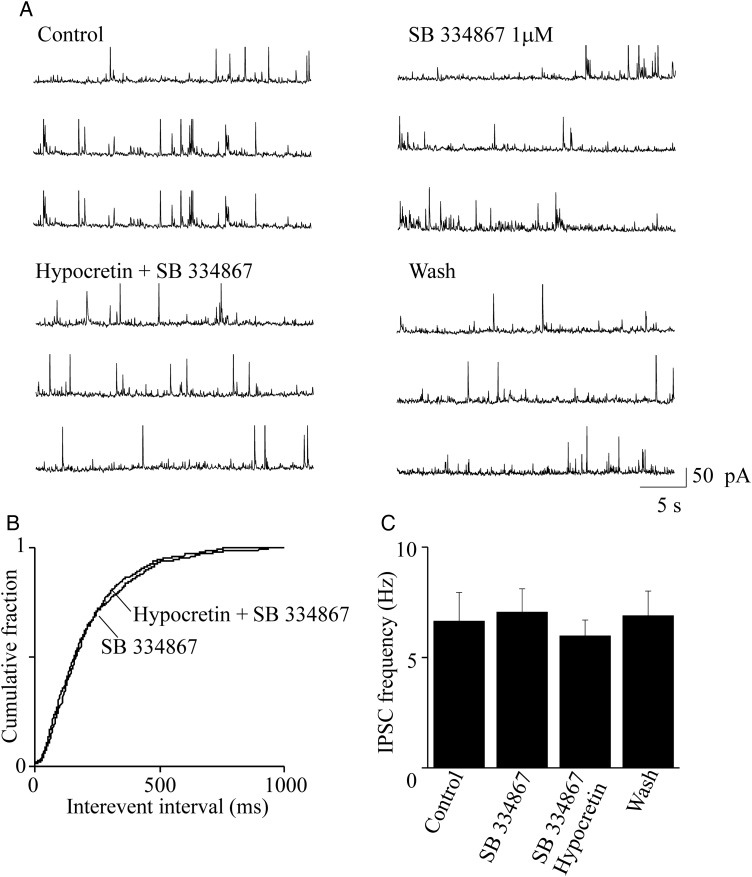
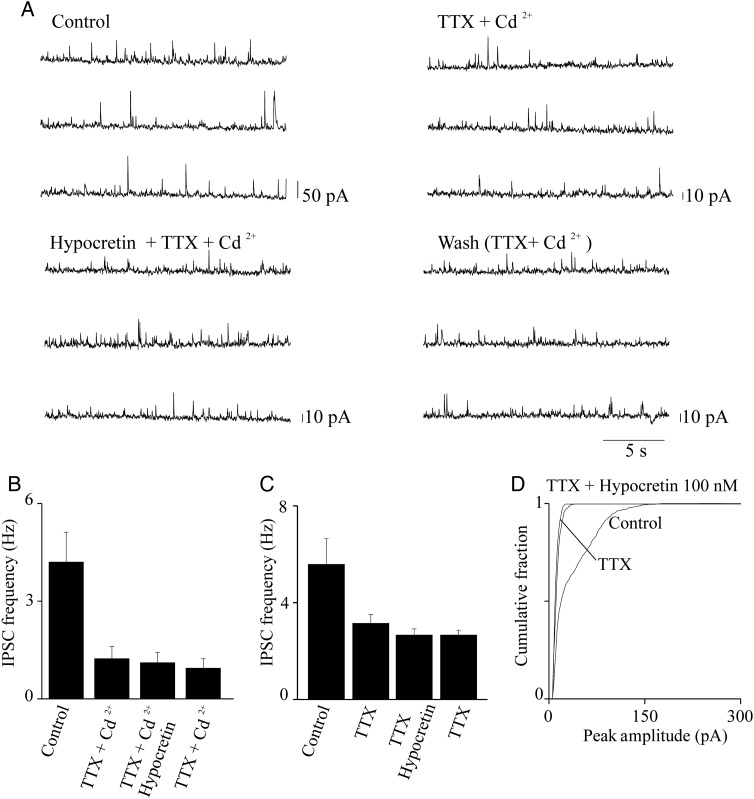
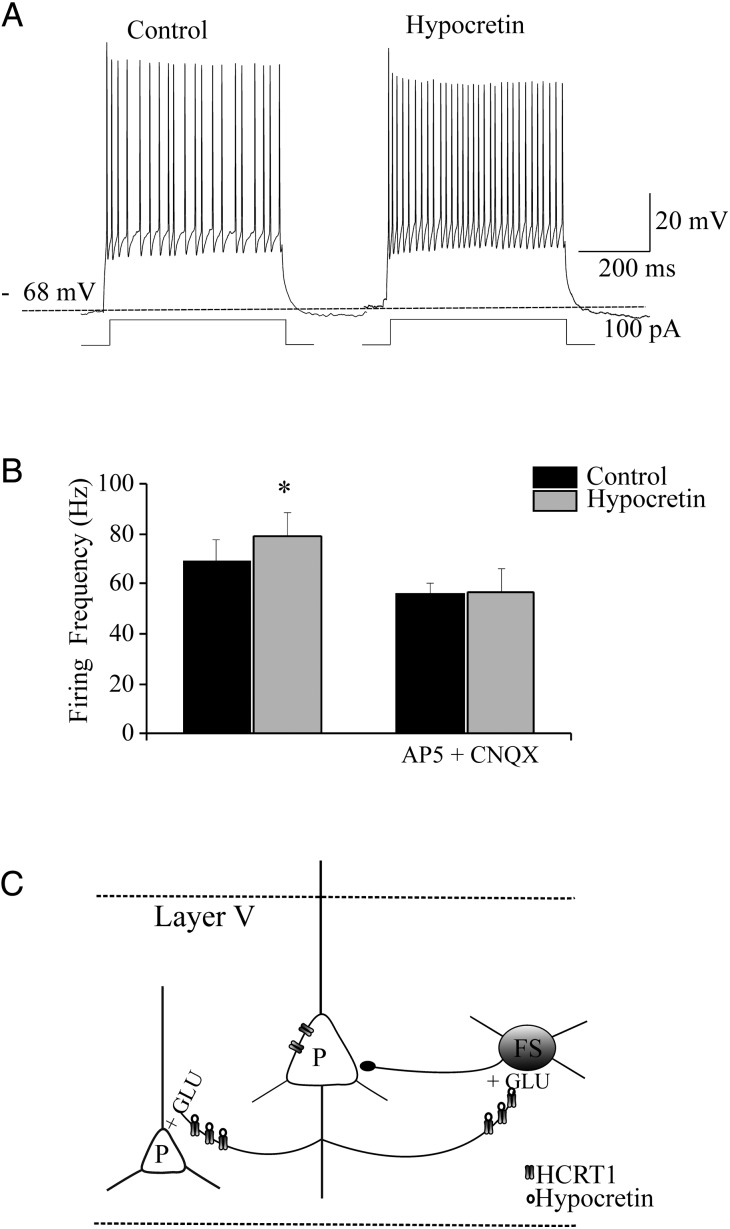
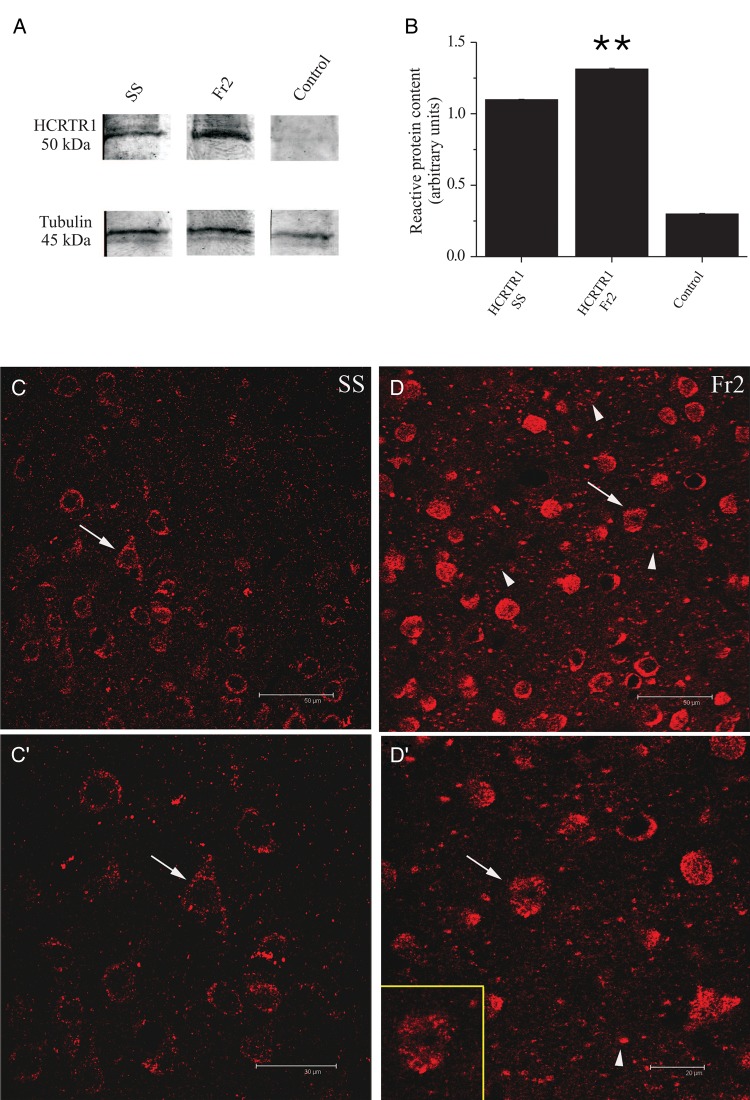
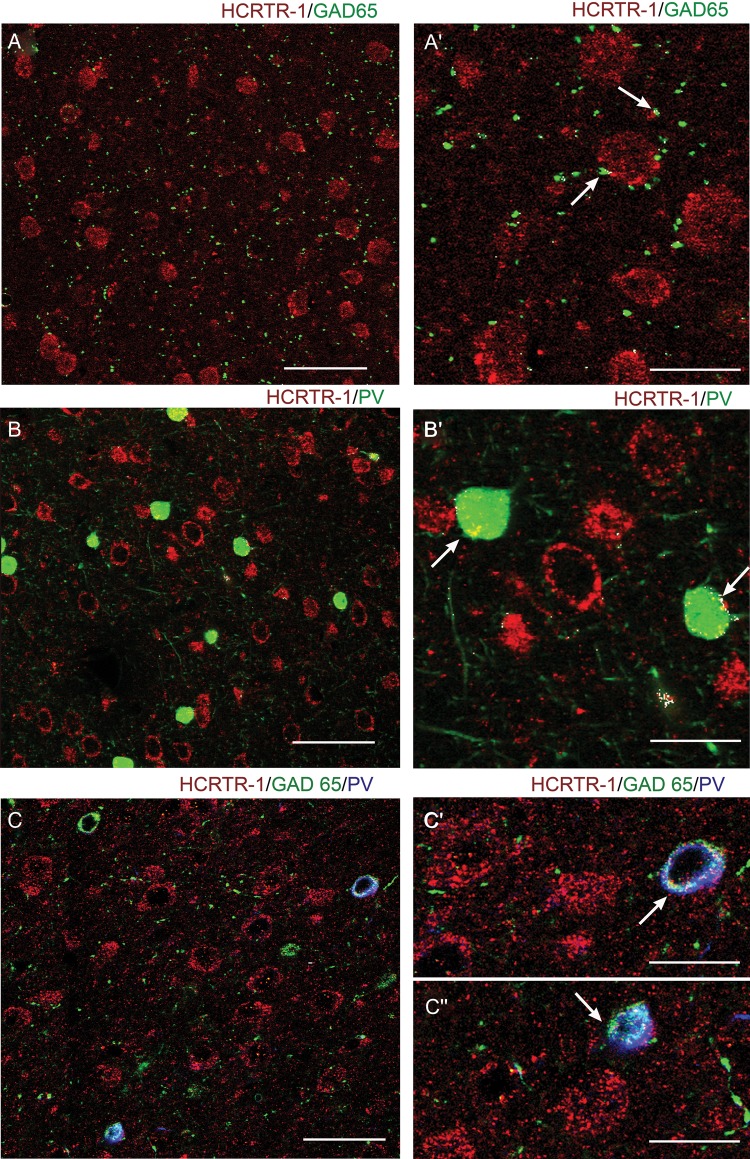
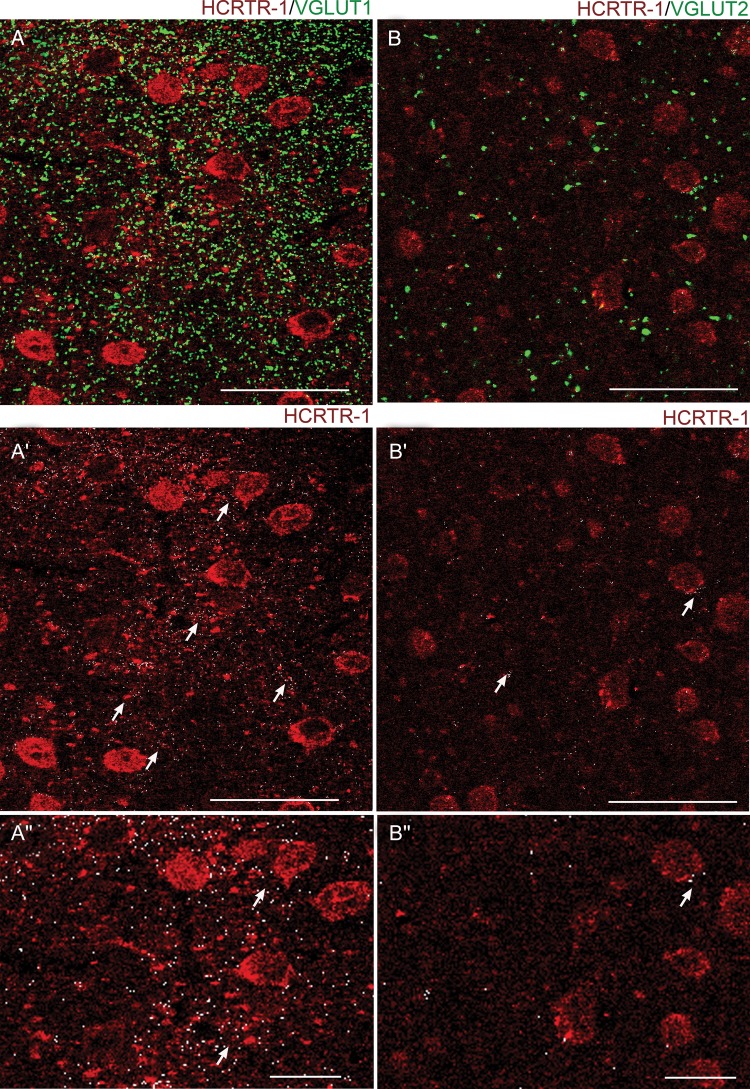
We studied the effect of hypocretin 1 (orexin A) in the frontal area 2 (Fr2) of the murine neocortex, implicated in the motivation-dependent goal-directed tasks. In layer V, hypocretin stimulated the spontaneous excitatory postsynaptic currents (EPSCs) on fast-spiking (FS) interneurons. The effect was accompanied by increased frequency of miniature EPSCs, indicating that hypocretin can target the glutamatergic terminals. Moreover, hypocretin stimulated the spontaneous inhibitory postsynaptic currents (IPSCs) on pyramidal neurons, with no effect on miniature IPSCs. This action was prevented by blocking 1) the ionotropic glutamatergic receptors; 2) the hypocretin receptor type 1 (HCRTR-1), with SB-334867. Finally, hypocretin increased the firing frequency in FS cells, and the effect was blocked when the ionotropic glutamate transmission was inhibited. Immunolocalization confirmed that HCRTR-1 is highly expressed in Fr2, particularly in layer V-VI. Conspicuous labeling was observed in pyramidal neuron somata and in VGLUT1+ glutamatergic terminals, but not in VGLUT2+ fibers (mainly thalamocortical afferents). The expression of HCRTR-1 in GABAergic structures was scarce. We conclude that 1) hypocretin regulates glutamate release in Fr2; 2) the effect presents a presynaptic component; 3) the peptide control of FS cells is indirect, and probably mediated by the regulation of glutamatergic input onto these cells.
Keywords: HCRTR; OXR1; SB-334867; VGLUT; fast-spiking; premotor.
© The Author 2013. Published by Oxford University Press.
Figures
References
-
- Ammoun S, Holmqvist T, Shariatmadari R, Oonk HB, Detheux M, Parmentier M, Akerman KE, Kukkonen JP. 2003. Distinct recognition of OX1 and OX2 receptors by orexin peptides. J Pharmacol Exp Ther. 305:507–514. - PubMed
-
- Aracri P, Amadeo A, Pasini ME, Fascio U, Becchetti A. 2013. Regulation of glutamate release by heteromeric nicotinic receptors in layer V of the secondary motor region (Fr2) in the dorsomedial shoulder of prefrontal cortex in mouse. Synapse. 67:338–357. - PubMed
-
- Aracri P, Consonni S, Morini R, Perrella M, Rodighiero S, Amadeo A, Becchetti A. 2010. Tonic modulation of GABA release by nicotinic acetylcholine receptors, in layer V of the murine prefrontal cortex. Cereb Cortex. 20:1539–1555. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
