

Cellular distribution of copper to superoxide dismutase involves scaffolding by membranes
- PMID: 24297923
- PMCID: PMC3870662
- DOI: 10.1073/pnas.1309820110
Cellular distribution of copper to superoxide dismutase involves scaffolding by membranes
Abstract
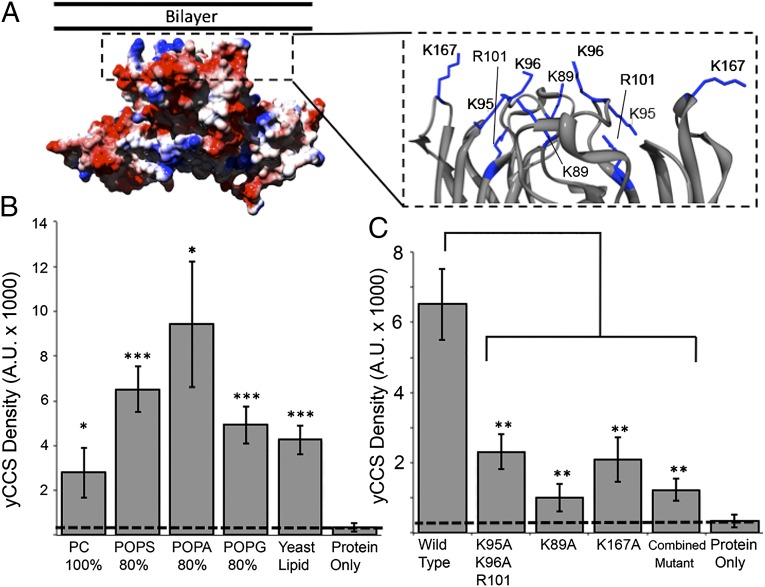
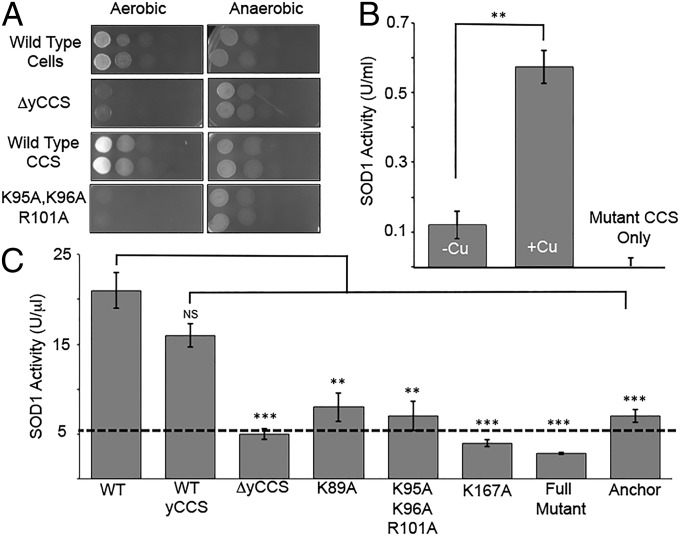
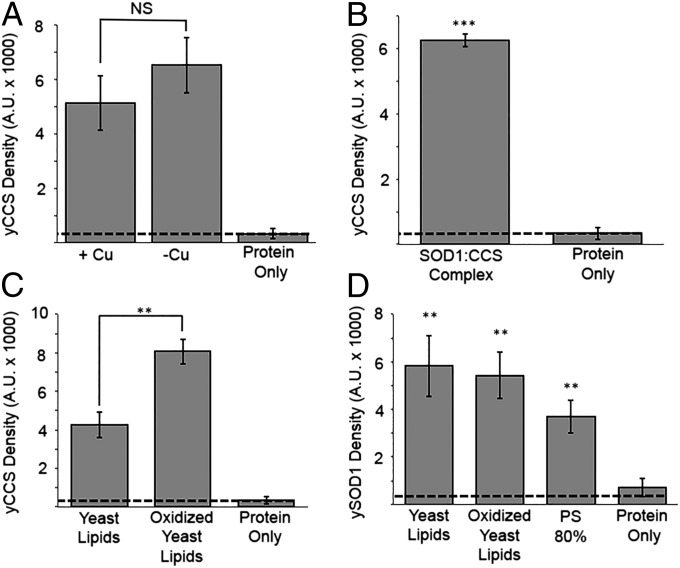
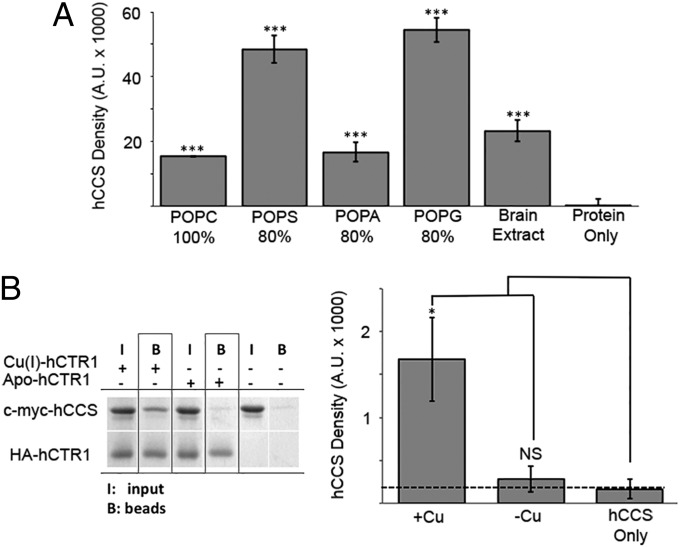
Efficient delivery of copper ions to specific intracellular targets requires copper chaperones that acquire metal cargo through unknown mechanisms. Here we demonstrate that the human and yeast copper chaperones (CCS) for superoxide dismutase 1 (SOD1), long thought to exclusively reside in the cytosol and mitochondrial intermembrane space, can engage negatively charged bilayers through a positively charged lipid-binding interface. The significance of this membrane-binding interface is established through SOD1 activity and genetic complementation studies in Saccharomyces cerevisiae, showing that recruitment of CCS to the membrane is required for activation of SOD1. Moreover, we show that a CCS:SOD1 complex binds to bilayers in vitro and that CCS can interact with human high affinity copper transporter 1. Shifting current paradigms, we propose that CCS-dependent copper acquisition and distribution largely occur at membrane interfaces and that this emerging role of the bilayer may reflect a general mechanistic aspect of cellular transition metal ion acquisition.
Keywords: copper homeostasis; copper transfer; copper uptake; membrane scaffold.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

Similar articles
-
Instability of superoxide dismutase 1 of Drosophila in mutants deficient for its cognate copper chaperone.J Biol Chem. 2008 Dec 19;283(51):35393-401. doi: 10.1074/jbc.M807131200. Epub 2008 Oct 23. J Biol Chem. 2008. PMID: 18948262 Free PMC article.
-
An essential role of N-terminal domain of copper chaperone in the enzymatic activation of Cu/Zn-superoxide dismutase.J Inorg Biochem. 2017 Oct;175:208-216. doi: 10.1016/j.jinorgbio.2017.07.036. Epub 2017 Jul 31. J Inorg Biochem. 2017. PMID: 28780408
-
Transcriptional activation in yeast in response to copper deficiency involves copper-zinc superoxide dismutase.J Biol Chem. 2009 Jan 2;284(1):404-413. doi: 10.1074/jbc.M807027200. Epub 2008 Oct 31. J Biol Chem. 2009. PMID: 18977757 Free PMC article.
-
Copper chaperones: personal escorts for metal ions.J Bioenerg Biomembr. 2002 Oct;34(5):373-9. doi: 10.1023/a:1021202119942. J Bioenerg Biomembr. 2002. PMID: 12539964 Review.
-
Import, maturation, and function of SOD1 and its copper chaperone CCS in the mitochondrial intermembrane space.Antioxid Redox Signal. 2010 Nov 1;13(9):1375-84. doi: 10.1089/ars.2010.3212. Antioxid Redox Signal. 2010. PMID: 20367259 Free PMC article. Review.
Cited by
-
Exploring the Extended Biological Functions of the Human Copper Chaperone of Superoxide Dismutase 1.Protein J. 2019 Aug;38(4):463-471. doi: 10.1007/s10930-019-09824-9. Protein J. 2019. PMID: 31140034 Review.
-
Superoxide Dismutase 1 in Health and Disease: How a Frontline Antioxidant Becomes Neurotoxic.Angew Chem Int Ed Engl. 2021 Apr 19;60(17):9215-9246. doi: 10.1002/anie.202000451. Epub 2020 Nov 19. Angew Chem Int Ed Engl. 2021. PMID: 32144830 Free PMC article. Review.
-
The TbD1 Locus Mediates a Hypoxia-Induced Copper Response in Mycobacterium bovis.Front Microbiol. 2022 Apr 14;13:817952. doi: 10.3389/fmicb.2022.817952. eCollection 2022. Front Microbiol. 2022. PMID: 35495699 Free PMC article.
-
Copper-zinc superoxide dismutase is activated through a sulfenic acid intermediate at a copper ion entry site.J Biol Chem. 2017 Jul 21;292(29):12025-12040. doi: 10.1074/jbc.M117.775981. Epub 2017 May 22. J Biol Chem. 2017. PMID: 28533431 Free PMC article.
-
Copper trafficking to the secretory pathway.Metallomics. 2016 Sep 1;8(9):840-52. doi: 10.1039/c6mt00176a. Epub 2016 Sep 5. Metallomics. 2016. PMID: 27603756 Free PMC article. Review.
References
-
- Bleackley MR, Macgillivray RT. Transition metal homeostasis: From yeast to human disease. Biometals. 2011;24(5):785–809. - PubMed
-
- Culotta V. Cell biology of copper. J Biol Inorg Chem. 2010;15(1):1–2. - PubMed
-
- Jomova K, Valko M. Advances in metal-induced oxidative stress and human disease. Toxicology. 2011;283(2-3):65–87. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
