

Independent RNA polymerase II preinitiation complex dynamics and nucleosome turnover at promoter sites in vivo
- PMID: 24298073
- PMCID: PMC3875852
- DOI: 10.1101/gr.157792.113
Independent RNA polymerase II preinitiation complex dynamics and nucleosome turnover at promoter sites in vivo
Abstract
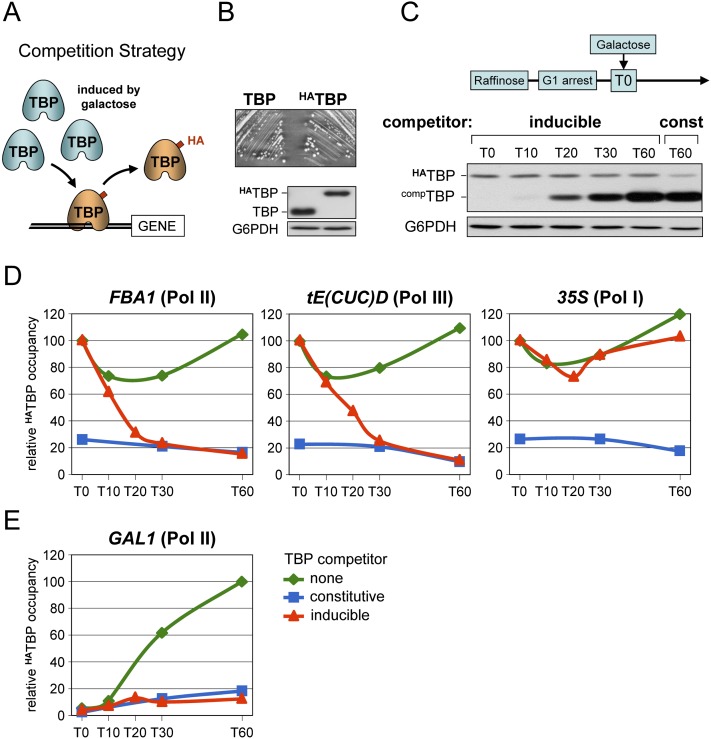
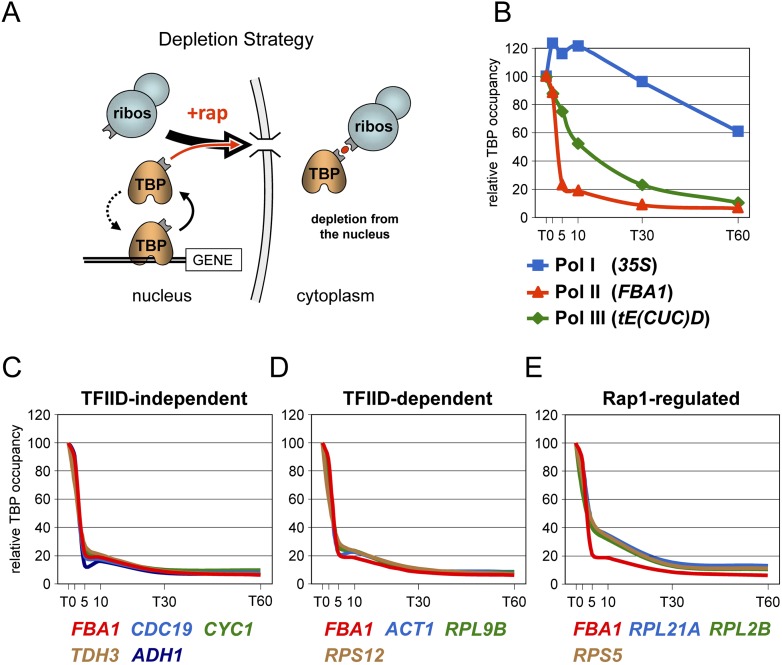
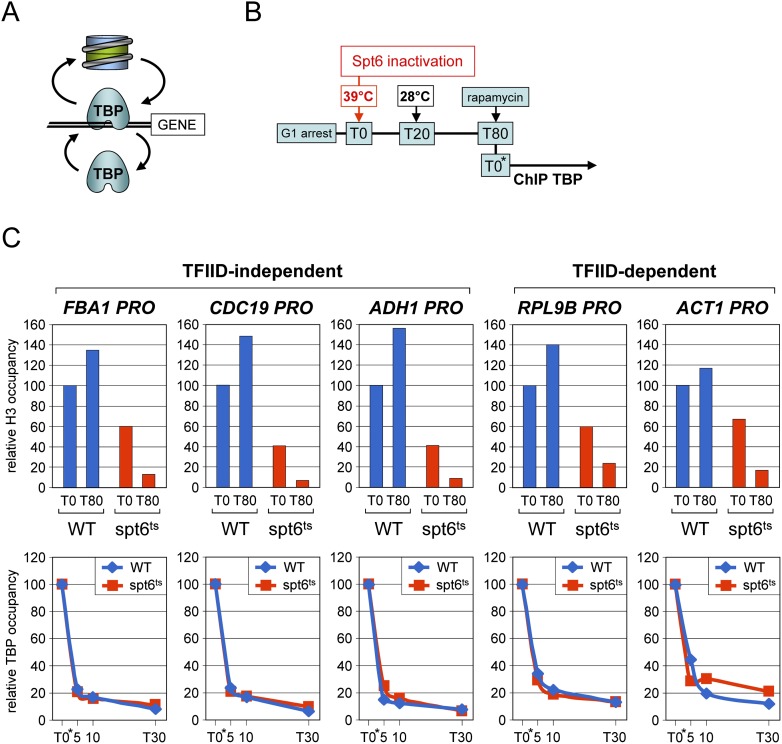
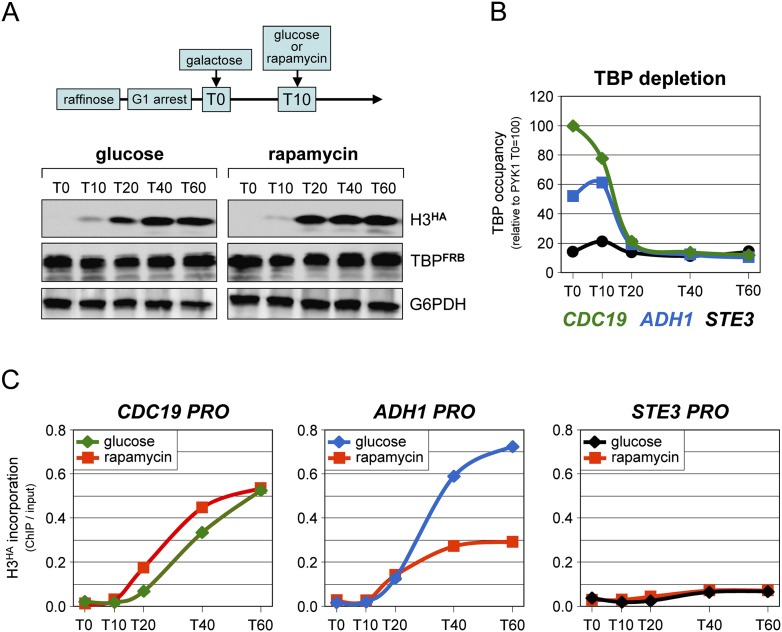
Transcription by all three eukaryotic RNA polymerases involves the assembly of a large preinitiation complex (PIC) at gene promoters. The PIC comprises several general transcription factors (GTFs), including TBP, and the respective RNA polymerase. It has been suggested that some GTFs remain stably bound at active promoters to facilitate multiple transcription events. Here we used two complementary approaches to show that, in G1-arrested yeast cells, TBP exchanges very rapidly even at the most highly active RNA Pol II promoters. A similar situation is observed at RNA Pol III promoters. In contrast, TBP remains stably bound at RNA Pol I promoters. We also provide evidence that, unexpectedly, PIC dynamics are neither the cause nor the consequence of nucleosome exchange at most of the RNA Pol II promoters we analyzed. These results point to a stable reinitiation complex at RNA Pol I promoters and suggest independent PIC and nucleosome turnover at many RNA Pol II promoters.
Figures
References
-
- Adkins MW, Tyler JK 2006. Transcriptional activators are dispensable for transcription in the absence of Spt6-mediated chromatin reassembly of promoter regions. Mol Cell 21: 405–416 - PubMed
-
- Bortvin A, Winston F 1996. Evidence that Spt6p controls chromatin structure by a direct interaction with histones. Science 272: 1473–1476 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases