

Zif268/Egr1 gain of function facilitates hippocampal synaptic plasticity and long-term spatial recognition memory
- PMID: 24298160
- PMCID: PMC3843890
- DOI: 10.1098/rstb.2013.0159
Zif268/Egr1 gain of function facilitates hippocampal synaptic plasticity and long-term spatial recognition memory
Abstract
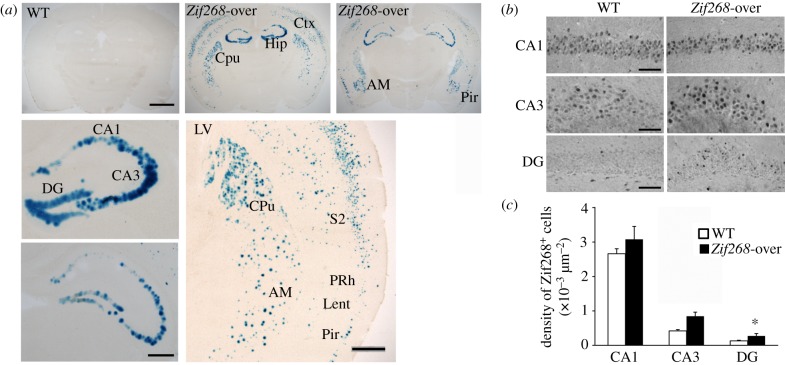
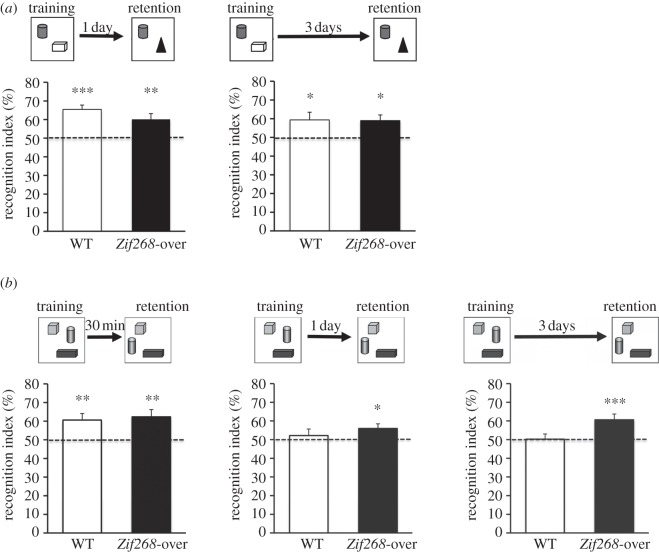
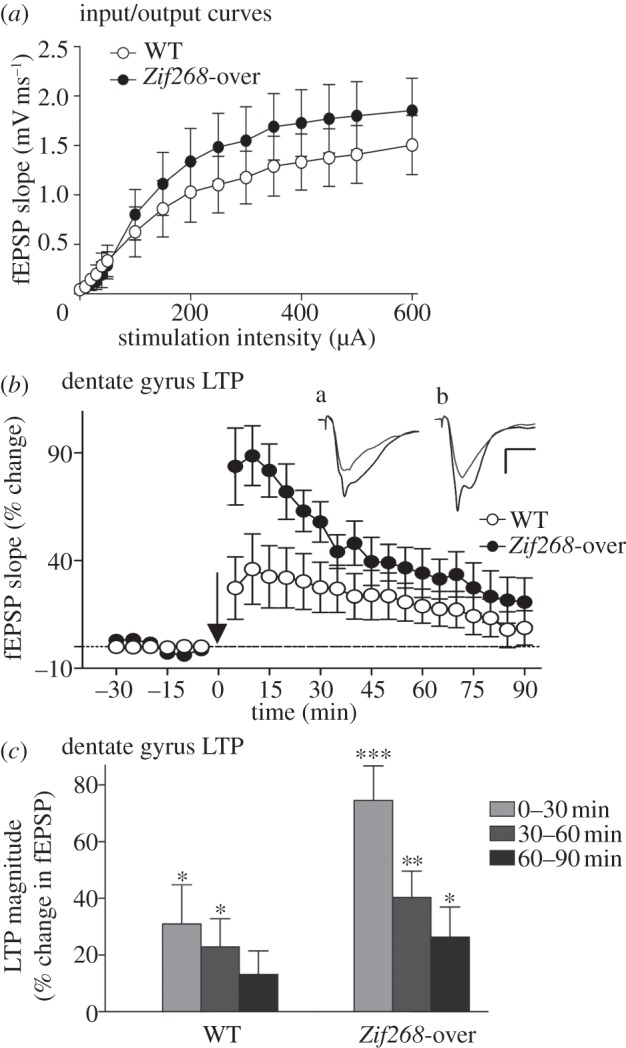
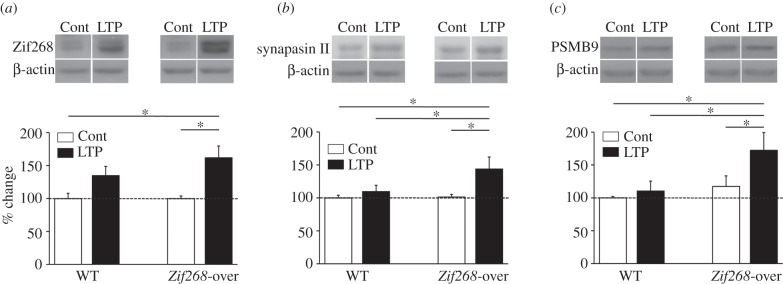
It is well established that Zif268/Egr1, a member of the Egr family of transcription factors, is critical for the consolidation of several forms of memory; however, it is as yet uncertain whether increasing expression of Zif268 in neurons can facilitate memory formation. Here, we used an inducible transgenic mouse model to specifically induce Zif268 overexpression in forebrain neurons and examined the effect on recognition memory and hippocampal synaptic transmission and plasticity. We found that Zif268 overexpression during the establishment of memory for objects did not change the ability to form a long-term memory of objects, but enhanced the capacity to form a long-term memory of the spatial location of objects. This enhancement was paralleled by increased long-term potentiation in the dentate gyrus of the hippocampus and by increased activity-dependent expression of Zif268 and selected Zif268 target genes. These results provide novel evidence that transcriptional mechanisms engaging Zif268 contribute to determining the strength of newly encoded memories.
Keywords: Egr1; conditional mutant mouse; dentate gyrus; long-term potentiation; recognition memory; transcription factor.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical