

How the mechanisms of long-term synaptic potentiation and depression serve experience-dependent plasticity in primary visual cortex
- PMID: 24298166
- PMCID: PMC3843896
- DOI: 10.1098/rstb.2013.0284
How the mechanisms of long-term synaptic potentiation and depression serve experience-dependent plasticity in primary visual cortex
Erratum in
- Philos Trans R Soc Lond B Biol Sci. 2014 Apr 5;369(1639):20140021
Abstract
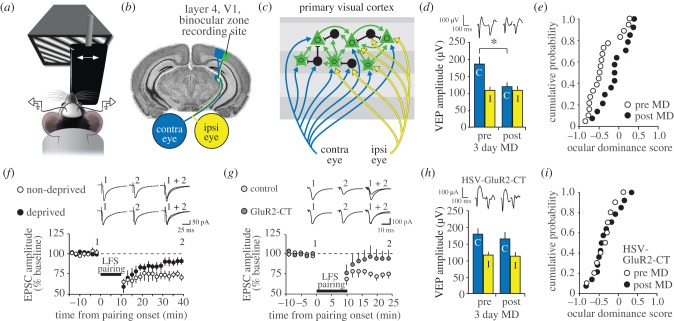
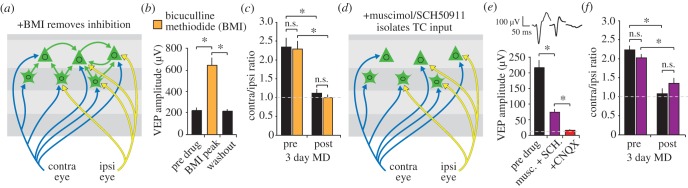
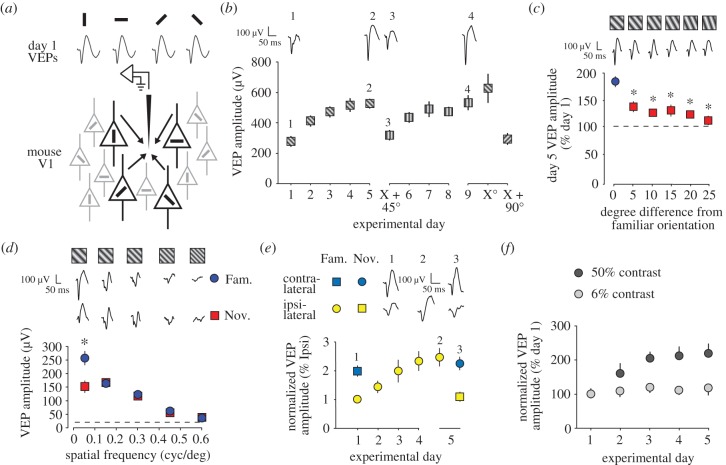
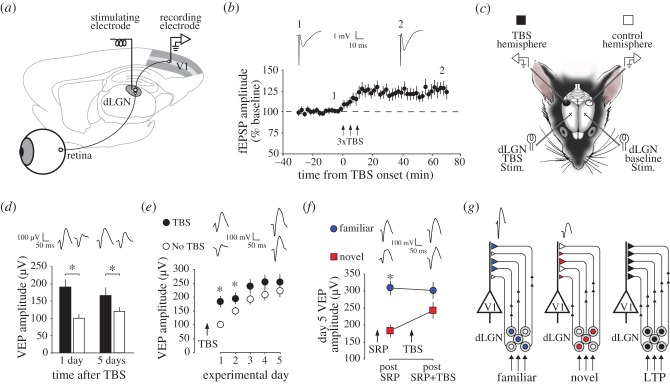
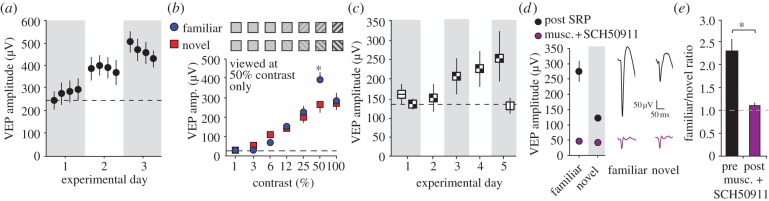
Donald Hebb chose visual learning in primary visual cortex (V1) of the rodent to exemplify his theories of how the brain stores information through long-lasting homosynaptic plasticity. Here, we revisit V1 to consider roles for bidirectional 'Hebbian' plasticity in the modification of vision through experience. First, we discuss the consequences of monocular deprivation (MD) in the mouse, which have been studied by many laboratories over many years, and the evidence that synaptic depression of excitatory input from the thalamus is a primary contributor to the loss of visual cortical responsiveness to stimuli viewed through the deprived eye. Second, we describe a less studied, but no less interesting form of plasticity in the visual cortex known as stimulus-selective response potentiation (SRP). SRP results in increases in the response of V1 to a visual stimulus through repeated viewing and bears all the hallmarks of perceptual learning. We describe evidence implicating an important role for potentiation of thalamo-cortical synapses in SRP. In addition, we present new data indicating that there are some features of this form of plasticity that cannot be fully accounted for by such feed-forward Hebbian plasticity, suggesting contributions from intra-cortical circuit components.
Keywords: amblyopia; long-term depression; long-term potentiation; monocular deprivation; perceptual learning; stimulus-selective response potentiation.
Figures
References
-
- Bain A. 1873. Mind and body. The theories of their relation. New York, NY: D. Appleton and Company.
-
- Hebb DO. 1949. The organization of behavior. New York, NY: Wiley.
-
- Wiesel TN, Hubel DH. 1963. Single-cell responses in striate cortex of kittens deprived of vision in one eye. J. Neurophysiol. 26, 1003–1017. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical