

Consequences of germline variation disrupting the constitutional translational initiation codon start sites of MLH1 and BRCA2: Use of potential alternative start sites and implications for predicting variant pathogenicity
- PMID: 24302565
- PMCID: PMC4041856
- DOI: 10.1002/mc.22116
Consequences of germline variation disrupting the constitutional translational initiation codon start sites of MLH1 and BRCA2: Use of potential alternative start sites and implications for predicting variant pathogenicity
Abstract
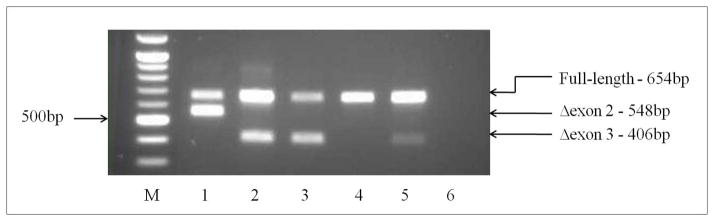
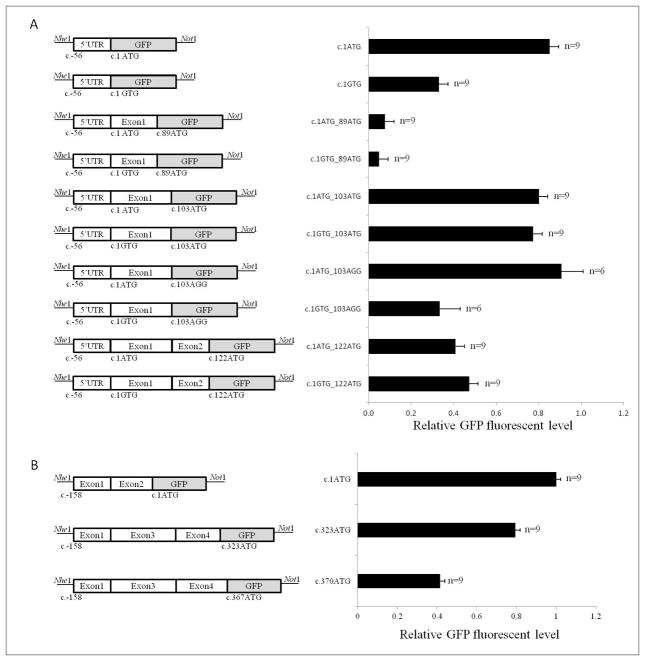
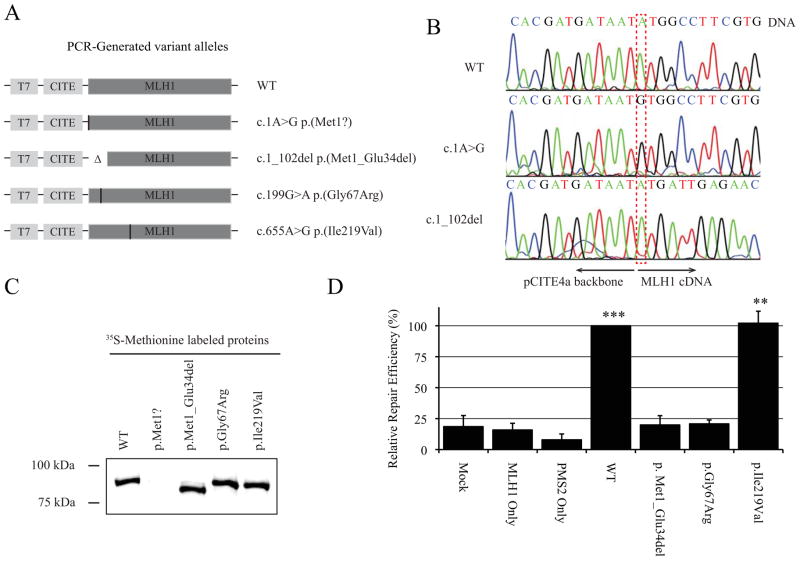
Variants that disrupt the translation initiation sequences in cancer predisposition genes are generally assumed to be deleterious. However, few studies have validated these assumptions with functional and clinical data. Two cancer syndrome gene variants likely to affect native translation initiation were identified by clinical genetic testing: MLH1:c.1A>G p.(Met1?) and BRCA2:c.67+3A>G. In vitro GFP-reporter assays were conducted to assess the consequences of translation initiation disruption on alternative downstream initiation codon usage. Analysis of MLH1:c.1A>G p.(Met1?) showed that translation was mostly initiated at an in-frame position 103 nucleotides downstream, but also at two ATG sequences downstream. The protein product encoded by the in-frame transcript initiating from position c.103 showed loss of in vitro mismatch repair activity comparable to known pathogenic mutations. BRCA2:c.67+3A>G was shown by mRNA analysis to result in an aberrantly spliced transcript deleting exon 2 and the consensus ATG site. In the absence of exon 2, translation initiated mostly at an out-of-frame ATG 323 nucleotides downstream, and to a lesser extent at an in-frame ATG 370 nucleotides downstream. Initiation from any of the downstream alternative sites tested in both genes would lead to loss of protein function, but further clinical data is required to confirm if these variants are associated with a high cancer risk. Importantly, our results highlight the need for caution in interpreting the functional and clinical consequences of variation that leads to disruption of the initiation codon, since translation may not necessarily occur from the first downstream alternative start site, or from a single alternative start site.
Keywords: cancer syndrome genes; in vitro assay; unclassified variant.
© 2013 Wiley Periodicals, Inc.
Figures

Similar articles
-
The importance of proper bioinformatics analysis and clinical interpretation of tumor genomic profiling: a case study of undifferentiated sarcoma and a constitutional pathogenic BRCA2 mutation and an MLH1 variant of uncertain significance.Fam Cancer. 2015 Sep;14(3):481-5. doi: 10.1007/s10689-015-9790-3. Fam Cancer. 2015. PMID: 25712765 Free PMC article.
-
Aberrant splicing in MLH1 and MSH2 due to exonic and intronic variants.Hum Genet. 2006 Mar;119(1-2):9-22. doi: 10.1007/s00439-005-0107-8. Epub 2005 Dec 8. Hum Genet. 2006. PMID: 16341550
-
Efficient use of a translation start codon in BDNF exon I.J Neurochem. 2015 Sep;134(6):1015-25. doi: 10.1111/jnc.13124. Epub 2015 Apr 27. J Neurochem. 2015. PMID: 25868795
-
A review of mismatch repair gene transcripts: issues for interpretation of mRNA splicing assays.Clin Genet. 2015 Feb;87(2):100-8. doi: 10.1111/cge.12450. Epub 2014 Jul 26. Clin Genet. 2015. PMID: 24989436 Review.
-
Novel approaches in evaluation of pathogenicity of single-base exonic germline changes involving the mismatch repair genes MLH1 and MSH2 in diagnostics of Lynch syndrome.Neoplasma. 2008;55(6):463-71. Neoplasma. 2008. PMID: 18999873 Review.
Cited by
-
Lynch syndrome mutation spectrum in New South Wales, Australia, including 55 novel mutations.Mol Genet Genomic Med. 2016 Jan 11;4(2):223-31. doi: 10.1002/mgg3.198. eCollection 2016 Mar. Mol Genet Genomic Med. 2016. PMID: 27064304 Free PMC article.
-
Biochemical and cellular consequences of the antithrombin p.Met1? mutation identified in a severe thrombophilic family.Oncotarget. 2018 Sep 4;9(69):33202-33214. doi: 10.18632/oncotarget.26059. eCollection 2018 Sep 4. Oncotarget. 2018. PMID: 30237862 Free PMC article.
-
In Silico Functional Annotation of Genomic Variation.Curr Protoc Hum Genet. 2016 Jan 1;88:6.15.1-6.15.17. doi: 10.1002/0471142905.hg0615s88. Curr Protoc Hum Genet. 2016. PMID: 26724722 Free PMC article. Review.
-
Adding In Silico Assessment of Potential Splice Aberration to the Integrated Evaluation of BRCA Gene Unclassified Variants.Hum Mutat. 2016 Jul;37(7):627-39. doi: 10.1002/humu.22973. Epub 2016 Apr 15. Hum Mutat. 2016. PMID: 26913838 Free PMC article.
-
Exome sequencing in genomic regions related to racing performance of Quarter Horses.J Appl Genet. 2019 Feb;60(1):79-86. doi: 10.1007/s13353-019-00483-1. Epub 2019 Jan 21. J Appl Genet. 2019. PMID: 30666567
References
-
- Parsons MT, Buchanan DD, Thompson B, Young JP, Spurdle AB. Correlation of tumour BRAF mutations and MLH1 methylation with germline mismatch repair (MMR) gene mutation status: a literature review assessing utility of tumour features for MMR variant classification. J Med Genet. 2012;49:151–7. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
