

The role of diacylglycerol kinase ζ and phosphatidic acid in the mechanical activation of mammalian target of rapamycin (mTOR) signaling and skeletal muscle hypertrophy
- PMID: 24302719
- PMCID: PMC3894336
- DOI: 10.1074/jbc.M113.531392
The role of diacylglycerol kinase ζ and phosphatidic acid in the mechanical activation of mammalian target of rapamycin (mTOR) signaling and skeletal muscle hypertrophy
Abstract
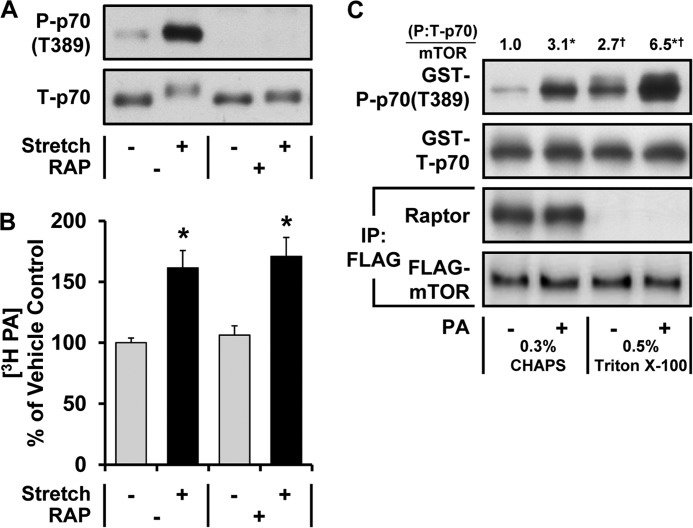
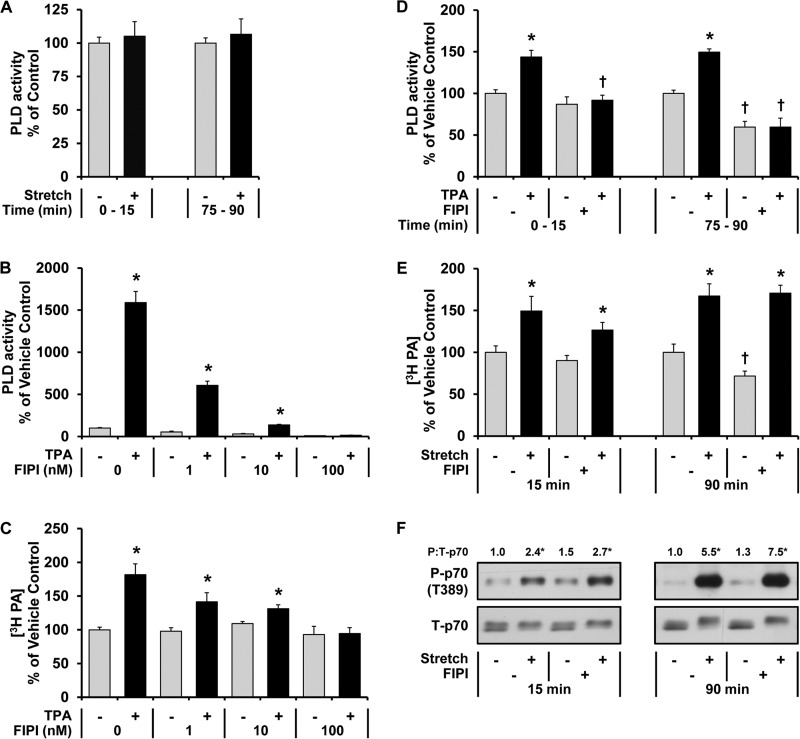
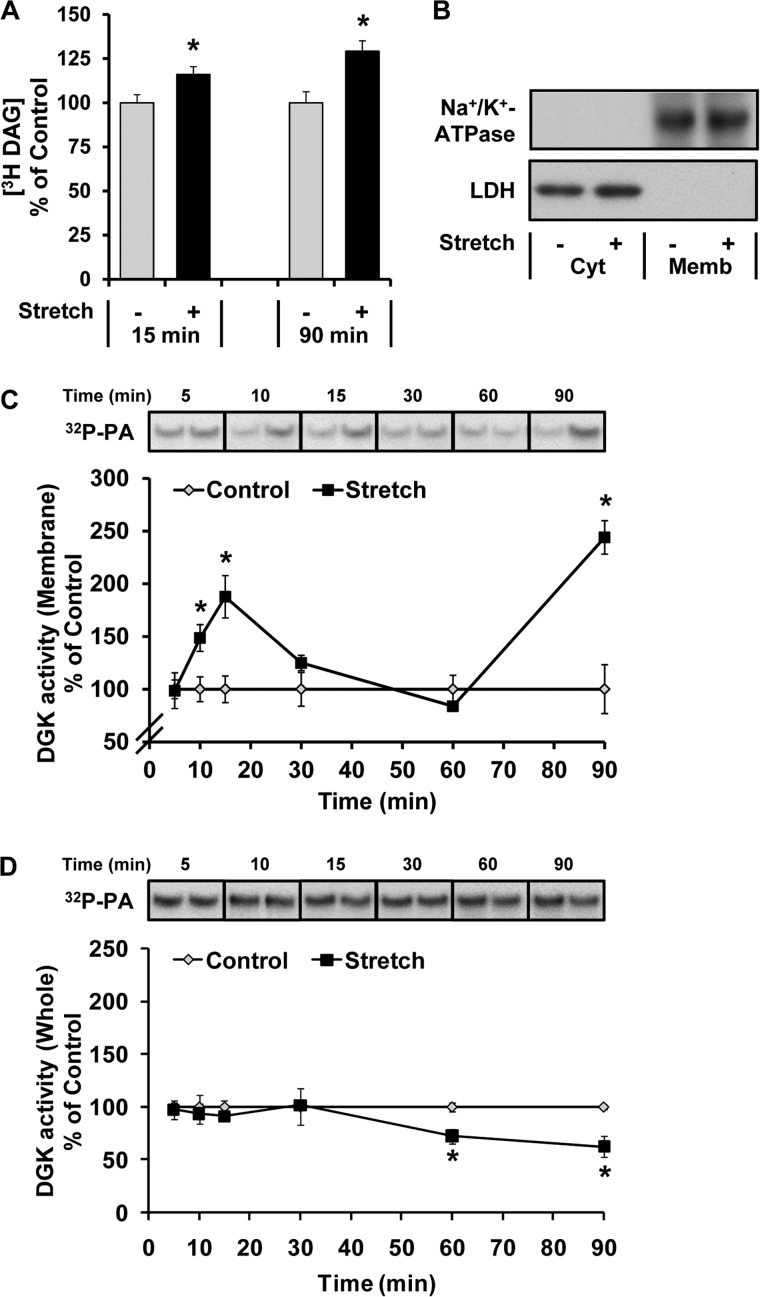
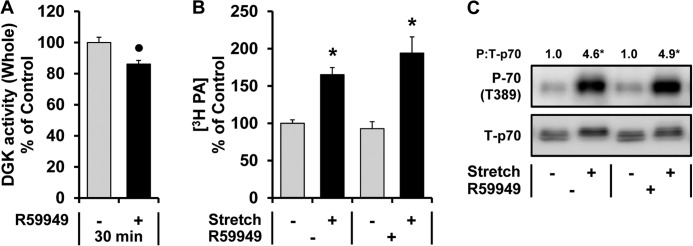
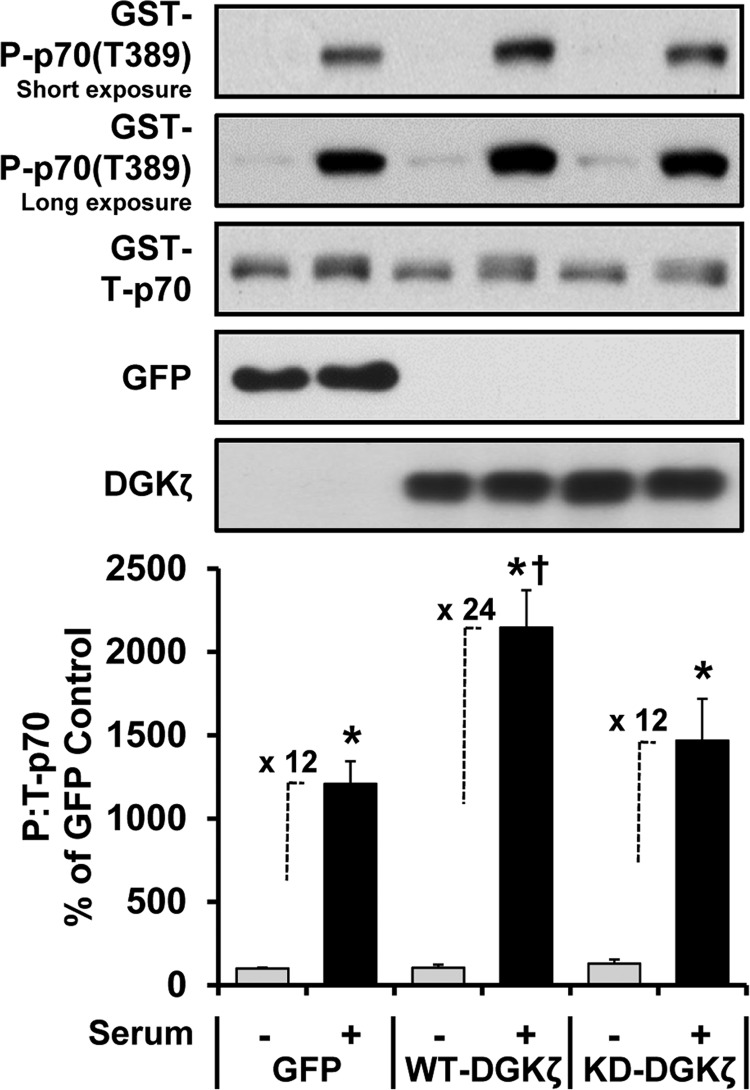
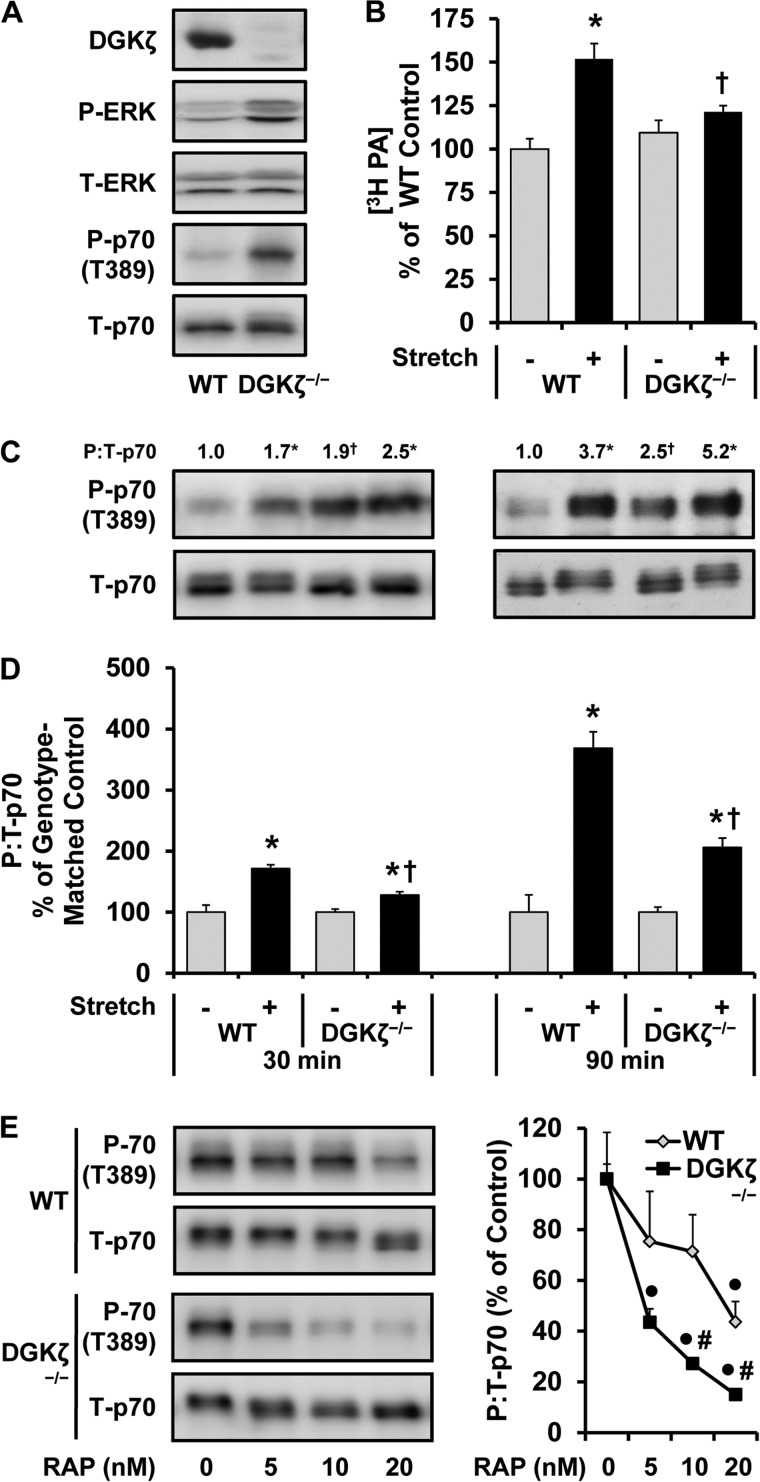
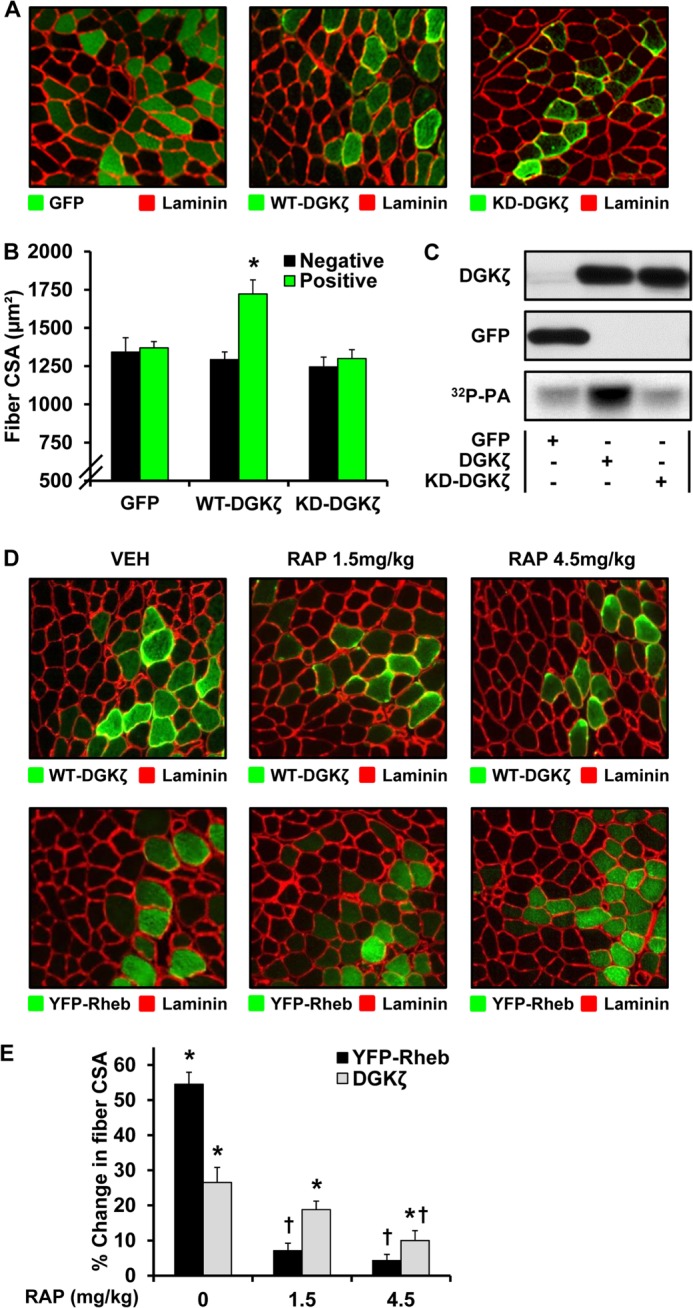
The activation of mTOR signaling is essential for mechanically induced changes in skeletal muscle mass, and previous studies have suggested that mechanical stimuli activate mTOR (mammalian target of rapamycin) signaling through a phospholipase D (PLD)-dependent increase in the concentration of phosphatidic acid (PA). Consistent with this conclusion, we obtained evidence which further suggests that mechanical stimuli utilize PA as a direct upstream activator of mTOR signaling. Unexpectedly though, we found that the activation of PLD is not necessary for the mechanically induced increases in PA or mTOR signaling. Motivated by this observation, we performed experiments that were aimed at identifying the enzyme(s) that promotes the increase in PA. These experiments revealed that mechanical stimulation increases the concentration of diacylglycerol (DAG) and the activity of DAG kinases (DGKs) in membranous structures. Furthermore, using knock-out mice, we determined that the ζ isoform of DGK (DGKζ) is necessary for the mechanically induced increase in PA. We also determined that DGKζ significantly contributes to the mechanical activation of mTOR signaling, and this is likely driven by an enhanced binding of PA to mTOR. Last, we found that the overexpression of DGKζ is sufficient to induce muscle fiber hypertrophy through an mTOR-dependent mechanism, and this event requires DGKζ kinase activity (i.e. the synthesis of PA). Combined, these results indicate that DGKζ, but not PLD, plays an important role in mechanically induced increases in PA and mTOR signaling. Furthermore, this study suggests that DGKζ could be a fundamental component of the mechanism(s) through which mechanical stimuli regulate skeletal muscle mass.
Keywords: Diacylglycerol Kinase ζ; Hypertrophy; Mechanotransduction; Phosphatidic Acid; Phospholipase D; Skeletal Muscle; mTOR.
Figures
References
-
- Hurley B. F., Hanson E. D., Sheaff A. K. (2011) Strength training as a countermeasure to aging muscle and chronic disease. Sports Med. 41, 289–306 - PubMed
-
- Goldberg A. L., Etlinger J. D., Goldspink D. F., Jablecki C. (1975) Mechanism of work-induced hypertrophy of skeletal muscle. Med. Sci. Sports 7, 185–198 - PubMed
-
- Avilés Mendoza G. J., Seidel N. E., Otsu M., Anderson S. M., Simon-Stoos K., Herrera A., Hoogstraten-Miller S., Malech H. L., Candotti F., Puck J. M., Bodine D. M. (2001) Comparison of five retrovirus vectors containing the human IL-2 receptor γ chain gene for their ability to restore T and B lymphocytes in the X-linked severe combined immunodeficiency mouse model. Mol. Ther. 3, 565–573 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
