

Dysferlin stabilizes stress-induced Ca2+ signaling in the transverse tubule membrane
- PMID: 24302765
- PMCID: PMC3870721
- DOI: 10.1073/pnas.1307960110
Dysferlin stabilizes stress-induced Ca2+ signaling in the transverse tubule membrane
Abstract
Dysferlinopathies, most commonly limb girdle muscular dystrophy 2B and Miyoshi myopathy, are degenerative myopathies caused by mutations in the DYSF gene encoding the protein dysferlin. Studies of dysferlin have focused on its role in the repair of the sarcolemma of skeletal muscle, but dysferlin's association with calcium (Ca(2+)) signaling proteins in the transverse (t-) tubules suggests additional roles. Here, we reveal that dysferlin is enriched in the t-tubule membrane of mature skeletal muscle fibers. Following experimental membrane stress in vitro, dysferlin-deficient muscle fibers undergo extensive functional and structural disruption of the t-tubules that is ameliorated by reducing external [Ca(2+)] or blocking L-type Ca(2+) channels with diltiazem. Furthermore, we demonstrate that diltiazem treatment of dysferlin-deficient mice significantly reduces eccentric contraction-induced t-tubule damage, inflammation, and necrosis, which resulted in a concomitant increase in postinjury functional recovery. Our discovery of dysferlin as a t-tubule protein that stabilizes stress-induced Ca(2+) signaling offers a therapeutic avenue for limb girdle muscular dystrophy 2B and Miyoshi myopathy patients.
Keywords: dihydropyridine receptor; excitation–contraction coupling; muscle injury; triad junction.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
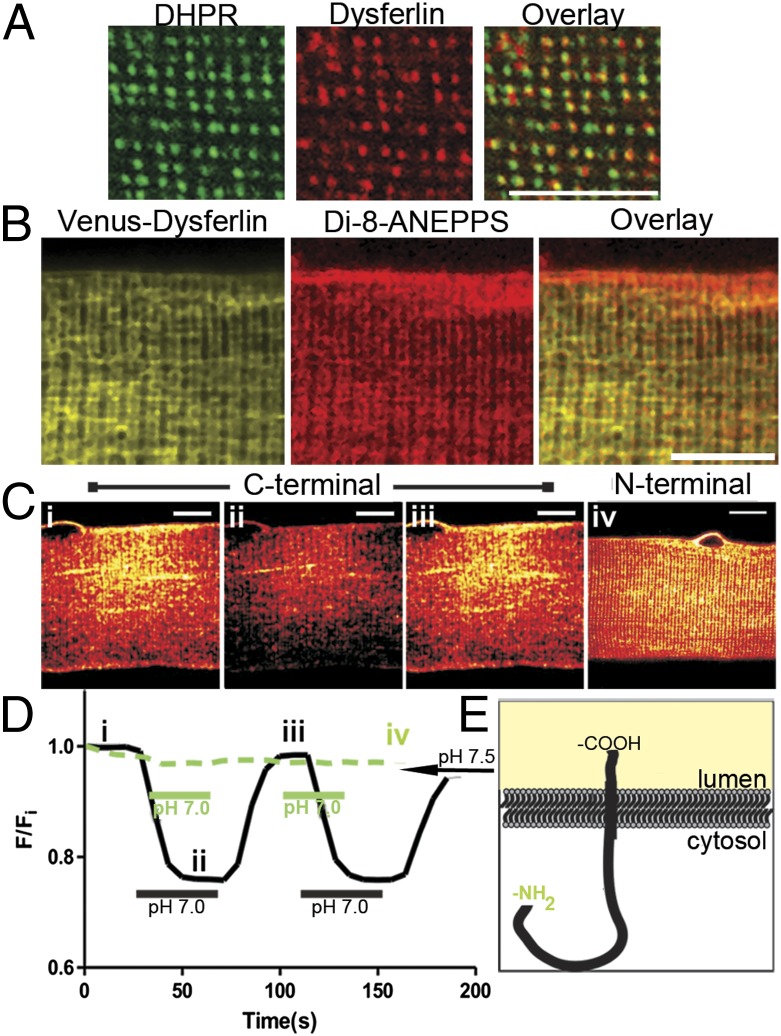
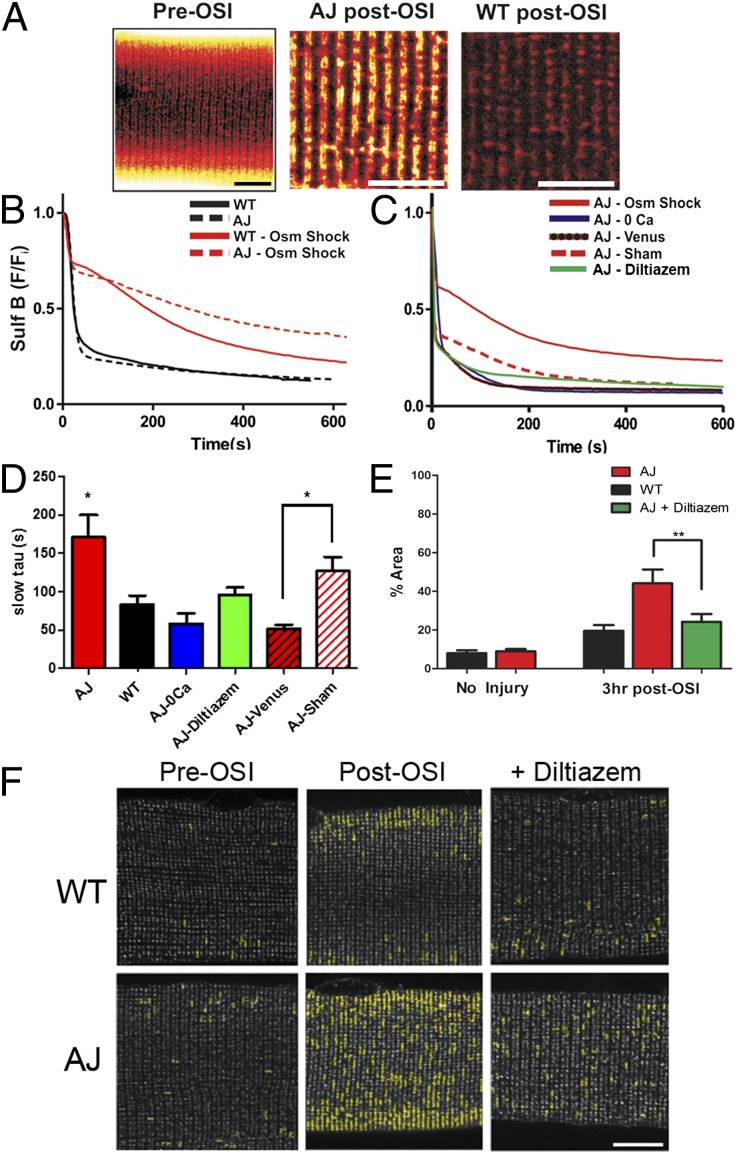
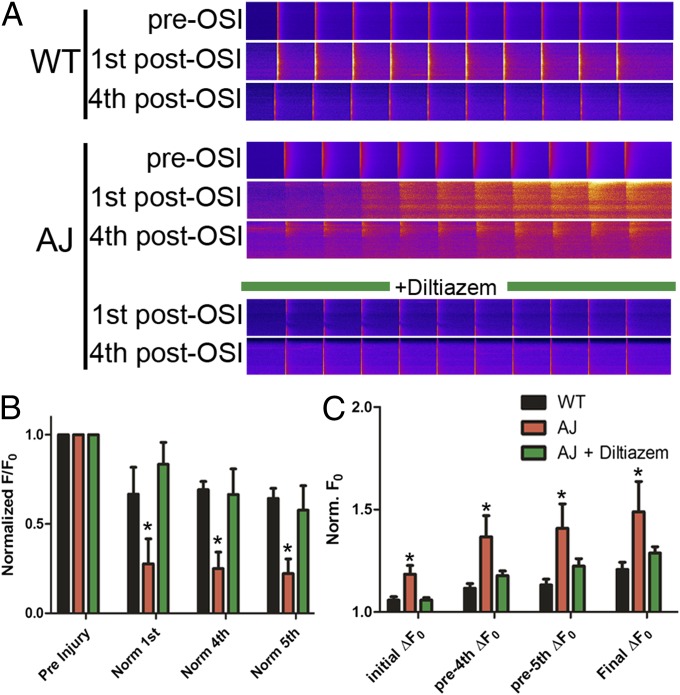
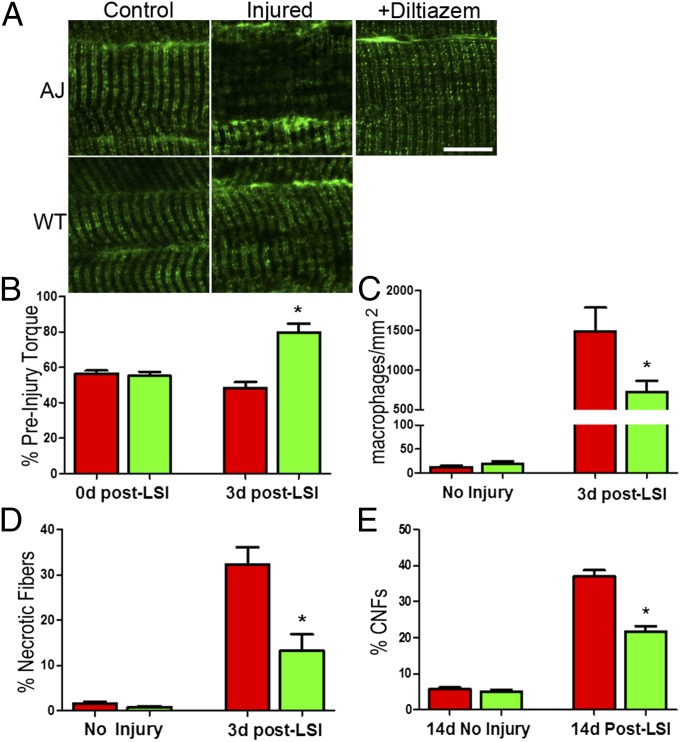
References
-
- Barohn RJ, Miller RG, Griggs RC. Autosomal recessive distal dystrophy. Neurology. 1991;41(9):1365–1370. - PubMed
-
- Bansal D, Campbell KP. Dysferlin and the plasma membrane repair in muscular dystrophy. Trends Cell Biol. 2004;14(4):206–213. - PubMed
-
- Bansal D, et al. Defective membrane repair in dysferlin-deficient muscular dystrophy. Nature. 2003;423(6936):168–172. - PubMed
-
- Lennon NJ, et al. Dysferlin interacts with annexins A1 and A2 and mediates sarcolemmal wound-healing. J Biol Chem. 2003;278(50):50466–50473. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
