

ANXUR receptor-like kinases coordinate cell wall integrity with growth at the pollen tube tip via NADPH oxidases
- PMID: 24302886
- PMCID: PMC3841104
- DOI: 10.1371/journal.pbio.1001719
ANXUR receptor-like kinases coordinate cell wall integrity with growth at the pollen tube tip via NADPH oxidases
Abstract
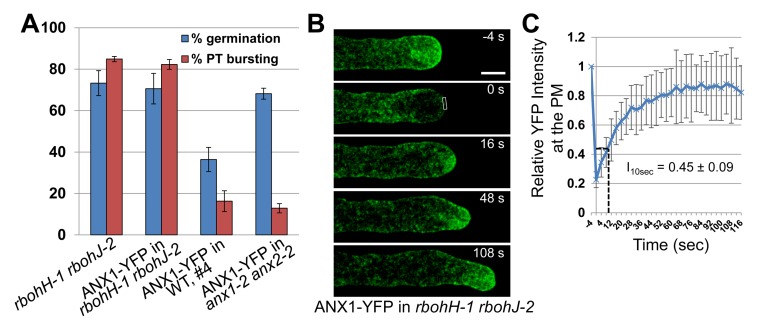
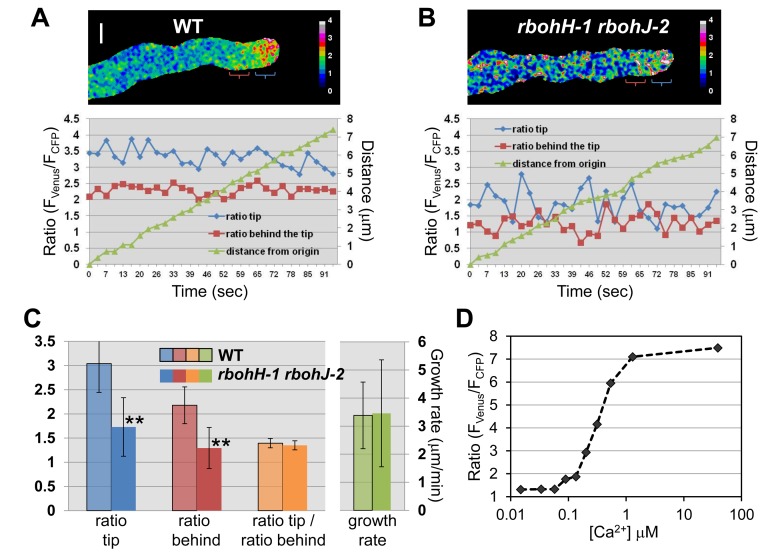
It has become increasingly apparent that the extracellular matrix (ECM), which in plants corresponds to the cell wall, can influence intracellular activities in ways that go far beyond their supposedly passive mechanical support. In plants, growing cells use mechanisms sensing cell wall integrity to coordinate cell wall performance with the internal growth machinery to avoid growth cessation or loss of integrity. How this coordination precisely works is unknown. Previously, we reported that in the tip-growing pollen tube the ANXUR receptor-like kinases (RLKs) of the CrRLK1L subfamily are essential to sustain growth without loss of cell wall integrity in Arabidopsis. Here, we show that over-expression of the ANXUR RLKs inhibits growth by over-activating exocytosis and the over-accumulation of secreted cell wall material. Moreover, the characterization of mutations in two partially redundant pollen-expressed NADPH oxidases coupled with genetic interaction studies demonstrate that the ANXUR RLKs function upstream of these NADPH oxidases. Using the H₂O₂-sensitive HyPer and the Ca²⁺-sensitive YC3.60 sensors in NADPH oxidase-deficient mutants, we reveal that NADPH oxidases generate tip-localized, pulsating H₂O₂ production that functions, possibly through Ca²⁺ channel activation, to maintain a steady tip-focused Ca²⁺ gradient during growth. Our findings support a model where ECM-sensing receptors regulate reactive oxygen species production, Ca²⁺ homeostasis, and exocytosis to coordinate ECM-performance with the internal growth machinery.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures





References
-
- Okayama H (2012) Cell cycle control by anchorage signaling. Cell Signal 24: 1599–1609. - PubMed
-
- Wolf S, Hématy K, Höfte H (2012) Growth control and cell wall signaling in plants. Annu Rev Plant Biol 63: 381–407. - PubMed
-
- Hématy K, Sado P-E, Van Tuinen A, Rochange S, Desnos T, et al. (2007) A receptor-like kinase mediates the response of Arabidopsis cells to the inhibition of cellulose synthesis. Curr Biol 17: 922–931. - PubMed
-
- Escobar-Restrepo J-M, Huck N, Kessler S, Gagliardini V, Gheyselinck J, et al. (2007) The FERONIA receptor-like kinase mediates male-female interactions during pollen tube reception. Science 317: 656–660. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
