

Respiratory syncytial virus induced type I IFN production by pDC is regulated by RSV-infected airway epithelial cells, RSV-exposed monocytes and virus specific antibodies
- PMID: 24303065
- PMCID: PMC3841124
- DOI: 10.1371/journal.pone.0081695
Respiratory syncytial virus induced type I IFN production by pDC is regulated by RSV-infected airway epithelial cells, RSV-exposed monocytes and virus specific antibodies
Abstract
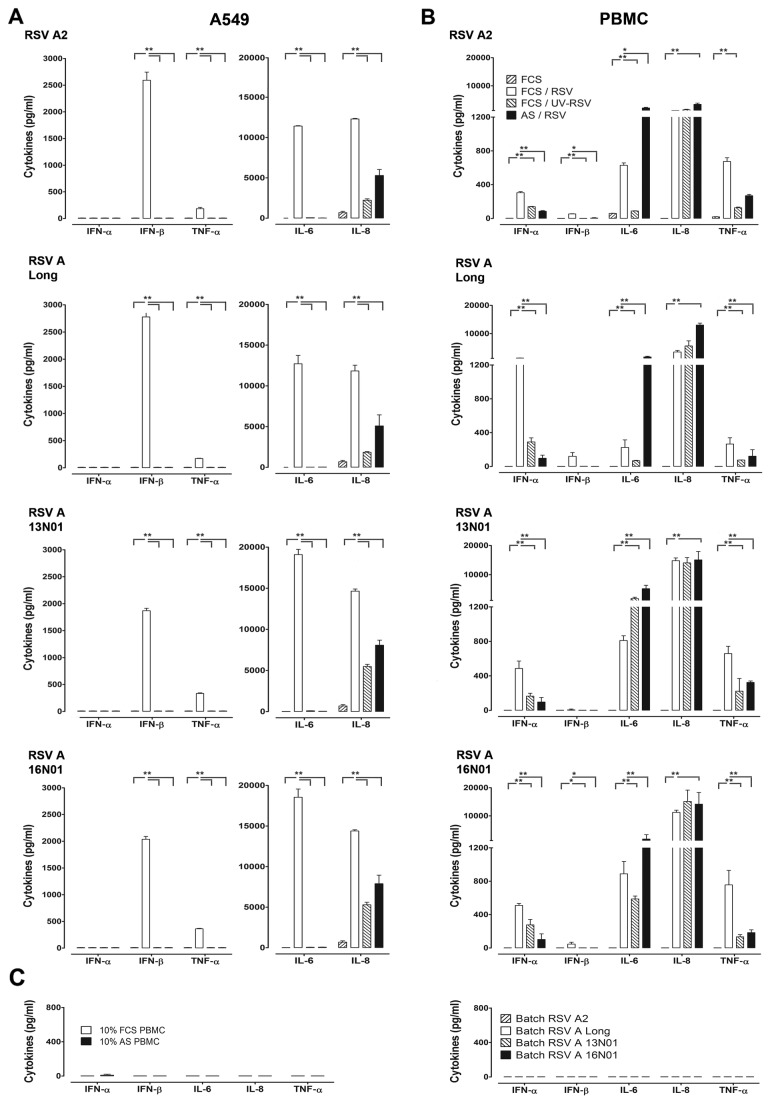
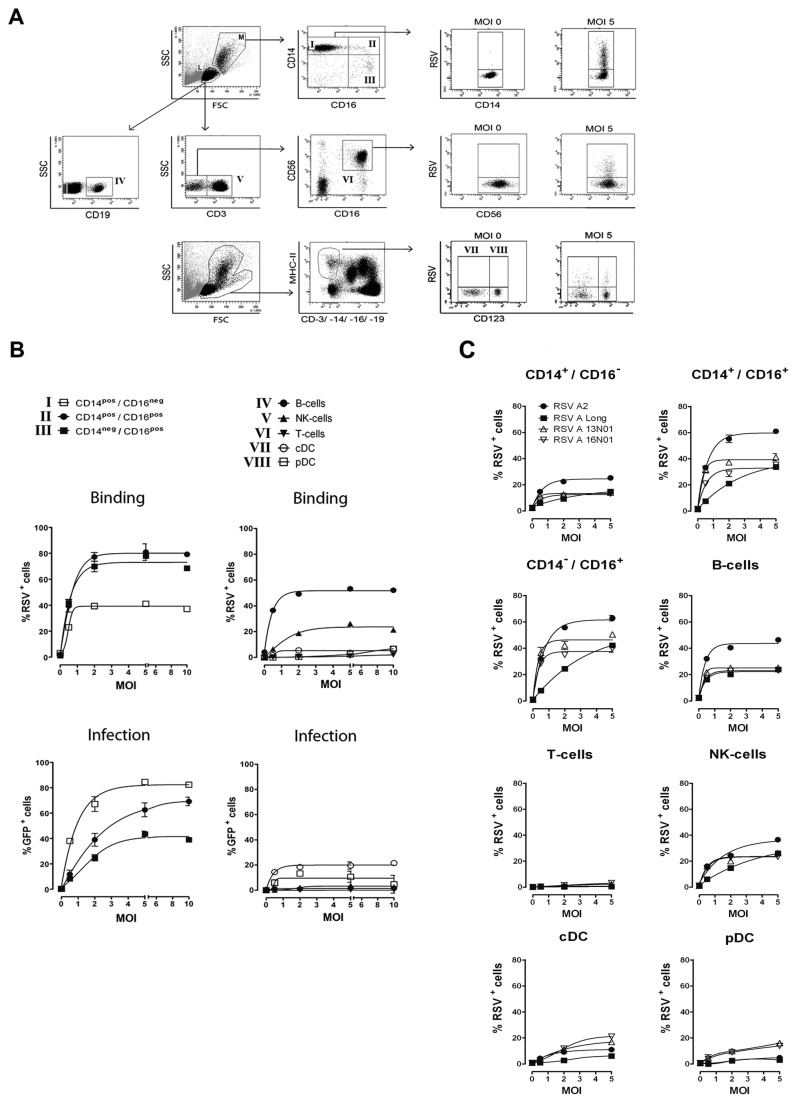
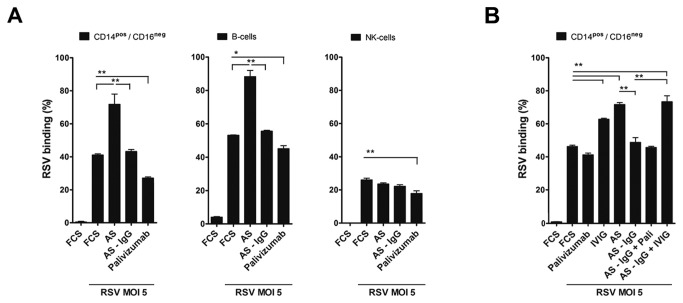
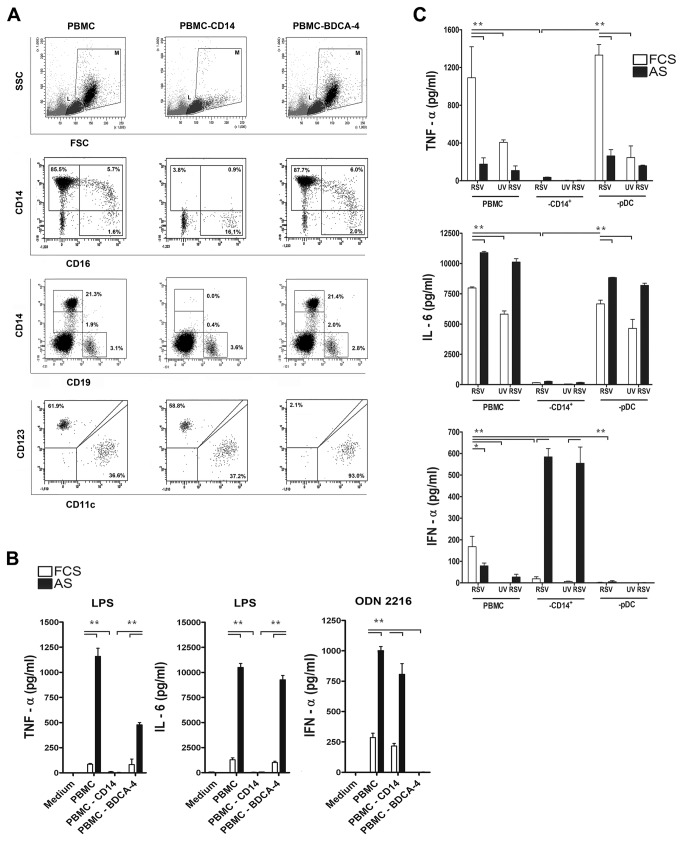
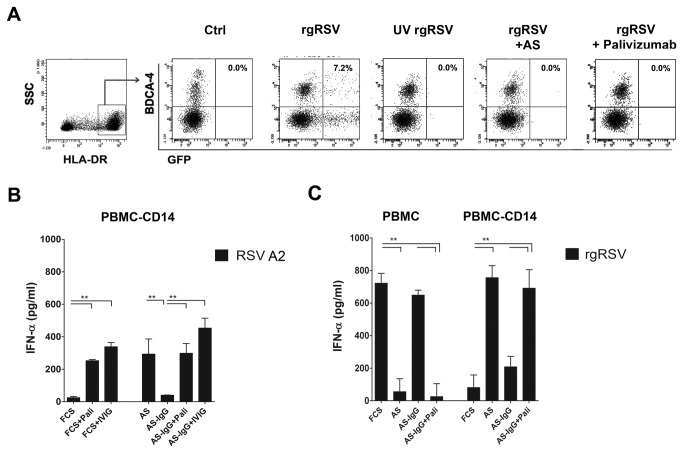
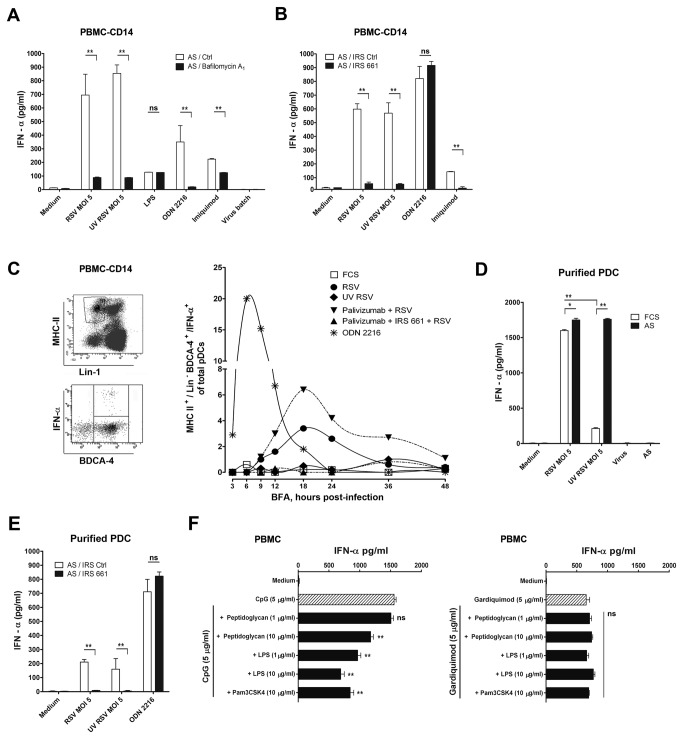
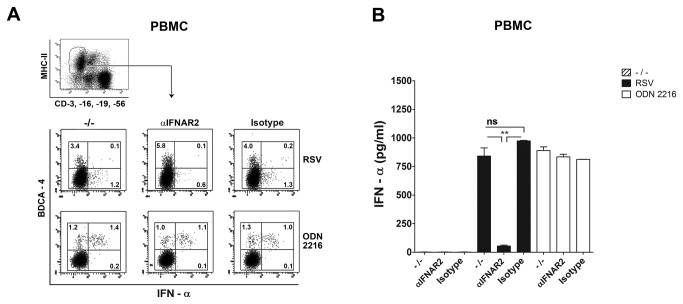
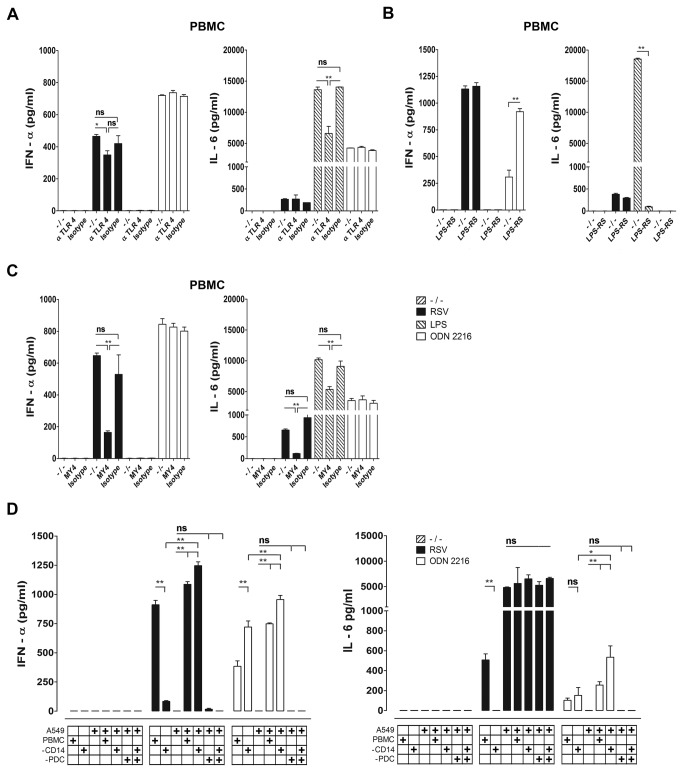
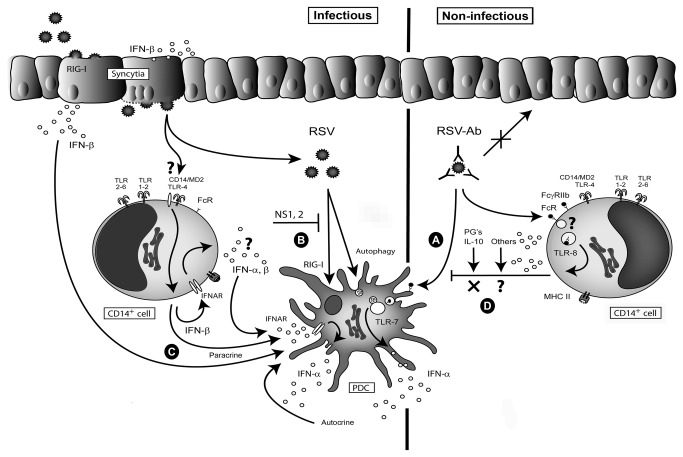
Innate immune responses elicited upon virus exposure are crucial for the effective eradication of viruses, the onset of adaptive immune responses and for establishing proper immune memory. Respiratory syncytial virus (RSV) is responsible for a high disease burden in neonates and immune compromised individuals, causing severe lower respiratory tract infections. During primary infections exuberant innate immune responses may contribute to disease severity. Furthermore, immune memory is often insufficient to protect during RSV re-exposure, which results in frequent symptomatic reinfections. Therefore, identifying the cell types and pattern recognition receptors (PRRs) involved in RSV-specific innate immune responses is necessary to understand incomplete immunity against RSV. We investigated the innate cellular response triggered upon infection of epithelial cells and peripheral blood mononuclear cells. We show that CD14(+) myeloid cells and epithelial cells are the major source of IL-8 and inflammatory cytokines, IL-6 and TNF-α, when exposed to live RSV Three routes of RSV-induced IFN-α production can be distinguished that depend on the cross-talk of different cell types and the presence or absence of virus specific antibodies, whereby pDC are the ultimate source of IFN-α. RSV-specific antibodies facilitate direct TLR7 access into endosomal compartments, while in the absence of antibodies, infection of monocytes or epithelial cells is necessary to provide an early source of type I interferons, required to engage the IFN-α,β receptor (IFNAR)-mediated pathway of IFN-α production by pDC. However, at high pDC density infection with RSV causes IFN-α production without the need for a second party cell. Our study shows that cellular context and immune status are factors affecting innate immune responses to RSV. These issues should therefore be addressed during the process of vaccine development and other interventions for RSV disease.
Conflict of interest statement
Figures
Similar articles
-
Attenuation of respiratory syncytial virus-induced and RIG-I-dependent type I IFN responses in human neonates and very young children.J Immunol. 2014 Feb 1;192(3):948-57. doi: 10.4049/jimmunol.1302007. Epub 2014 Jan 3. J Immunol. 2014. PMID: 24391215
-
Respiratory syncytial virus G protein CX3C motif impairs human airway epithelial and immune cell responses.J Virol. 2013 Dec;87(24):13466-79. doi: 10.1128/JVI.01741-13. Epub 2013 Oct 2. J Virol. 2013. PMID: 24089561 Free PMC article.
-
Platelets Modulate Innate Immune Response Against Human Respiratory Syncytial Virus In Vitro.Viral Immunol. 2017 Oct;30(8):576-581. doi: 10.1089/vim.2016.0161. Epub 2017 Aug 7. Viral Immunol. 2017. PMID: 28783457
-
How RSV Proteins Join Forces to Overcome the Host Innate Immune Response.Viruses. 2022 Feb 17;14(2):419. doi: 10.3390/v14020419. Viruses. 2022. PMID: 35216012 Free PMC article. Review.
-
Respiratory syncytial virus nonstructural proteins 1 and 2: Exceptional disrupters of innate immune responses.PLoS Pathog. 2019 Oct 17;15(10):e1007984. doi: 10.1371/journal.ppat.1007984. eCollection 2019 Oct. PLoS Pathog. 2019. PMID: 31622448 Free PMC article. Review.
Cited by
-
Contribution of Dendritic Cells in Protective Immunity against Respiratory Syncytial Virus Infection.Viruses. 2020 Jan 15;12(1):102. doi: 10.3390/v12010102. Viruses. 2020. PMID: 31952261 Free PMC article. Review.
-
Neutralizing antibodies inhibit HIV-1 infection of plasmacytoid dendritic cells by an FcγRIIa independent mechanism and do not diminish cytokines production.Sci Rep. 2014 Aug 18;4:5845. doi: 10.1038/srep05845. Sci Rep. 2014. PMID: 25132382 Free PMC article.
-
Interference Between Respiratory Syncytial Virus and Human Rhinovirus Infection in Infancy.J Infect Dis. 2017 Apr 1;215(7):1102-1106. doi: 10.1093/infdis/jix031. J Infect Dis. 2017. PMID: 28368456 Free PMC article. Clinical Trial.
-
Infection of nonclassic monocytes by respiratory syncytial virus induces an imbalance in the CD4+ T-cell subset response.Microbiol Spectr. 2025 Jan 7;13(1):e0207324. doi: 10.1128/spectrum.02073-24. Epub 2024 Dec 10. Microbiol Spectr. 2025. PMID: 39656009 Free PMC article.
-
Cellular RNA Helicase DDX1 Is Involved in Transmissible Gastroenteritis Virus nsp14-Induced Interferon-Beta Production.Front Immunol. 2017 Aug 9;8:940. doi: 10.3389/fimmu.2017.00940. eCollection 2017. Front Immunol. 2017. PMID: 28848548 Free PMC article.
References
-
- Janeway CA (1989) Approaching the asymptote? Evolution and revolution in immunology. Cold Spring Harbor Symposia on Quantitative Biology 54 1: 1–13. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
