

Connexin36 in gap junctions forming electrical synapses between motoneurons in sexually dimorphic motor nuclei in spinal cord of rat and mouse
- PMID: 24304165
- PMCID: PMC3943632
- DOI: 10.1111/ejn.12439
Connexin36 in gap junctions forming electrical synapses between motoneurons in sexually dimorphic motor nuclei in spinal cord of rat and mouse
Abstract
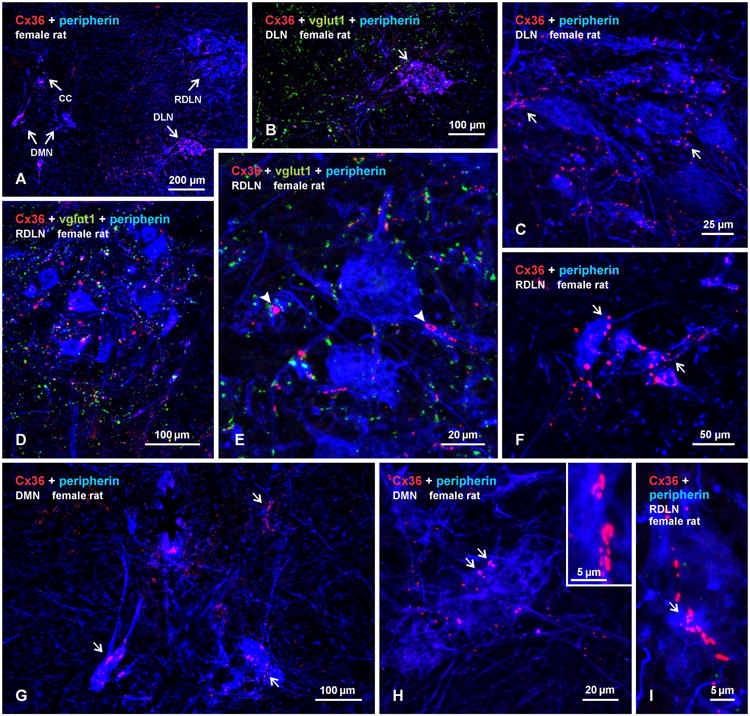
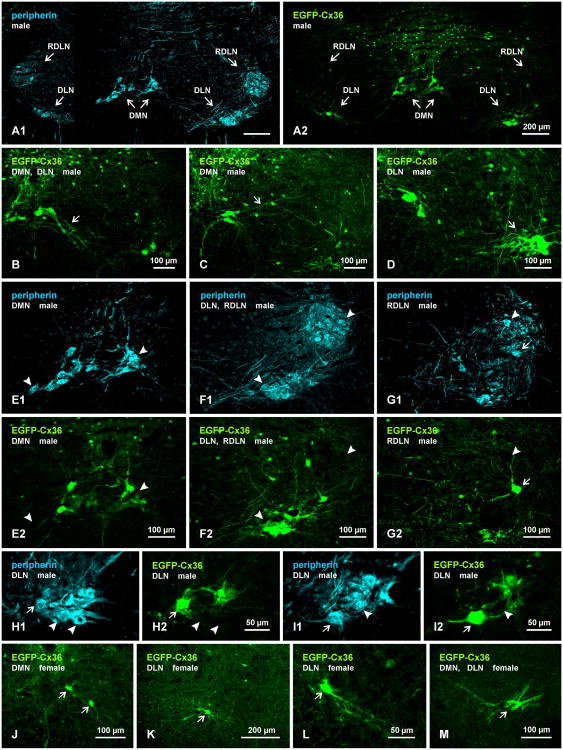
Pools of motoneurons in the lumbar spinal cord innervate the sexually dimorphic perineal musculature, and are themselves sexually dimorphic, showing differences in number and size between male and female rodents. In two of these pools, the dorsomedial nucleus (DMN) and the dorsolateral nucleus (DLN), dimorphic motoneurons are intermixed with non-dimorphic neurons innervating anal and external urethral sphincter muscles. As motoneurons in these nuclei are reportedly linked by gap junctions, we examined immunofluorescence labeling for the gap junction-forming protein connexin36 (Cx36) in male and female mice and rats. Fluorescent Cx36-labeled puncta occurred in distinctly greater amounts in the DMN and DLN of male rodents than in other spinal cord regions. These puncta were localized to motoneuron somata, proximal dendrites, and neuronal appositions, and were distributed either as isolated or large patches of puncta. In both rats and mice, Cx36-labeled puncta were associated with nearly all (> 94%) DMN and DLN motoneurons. The density of Cx36-labeled puncta increased dramatically from postnatal days 9 to 15, unlike the developmental decreases in these puncta observed in other central nervous system regions. In females, Cx36 labeling of puncta in the DLN was similar to that in males, but was sparse in the DMN. In enhanced green fluorescent protein (EGFP)-Cx36 transgenic mice, motoneurons in the DMN and DLN were intensely labeled for the EGFP reporter in males, but less so in females. The results indicate the presence of Cx36-containing gap junctions in the sexually dimorphic DMN and DLN of both male and female rodents, suggesting coupling of not only sexually dimorphic but also non-dimorphic motoneurons in these nuclei.
Keywords: EGFP-Cx36 reporter; gap junctions; immunofluorescence; motoneurons.
© 2013 Federation of European Neuroscience Societies and John Wiley & Sons Ltd.
Figures
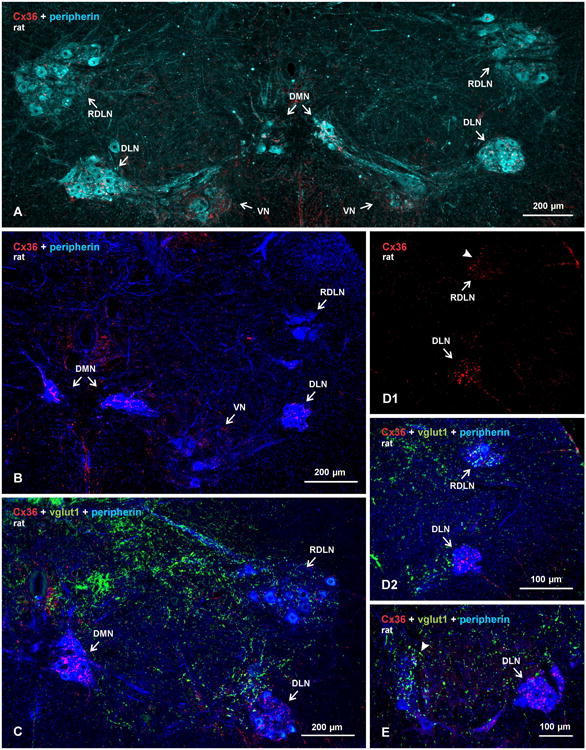
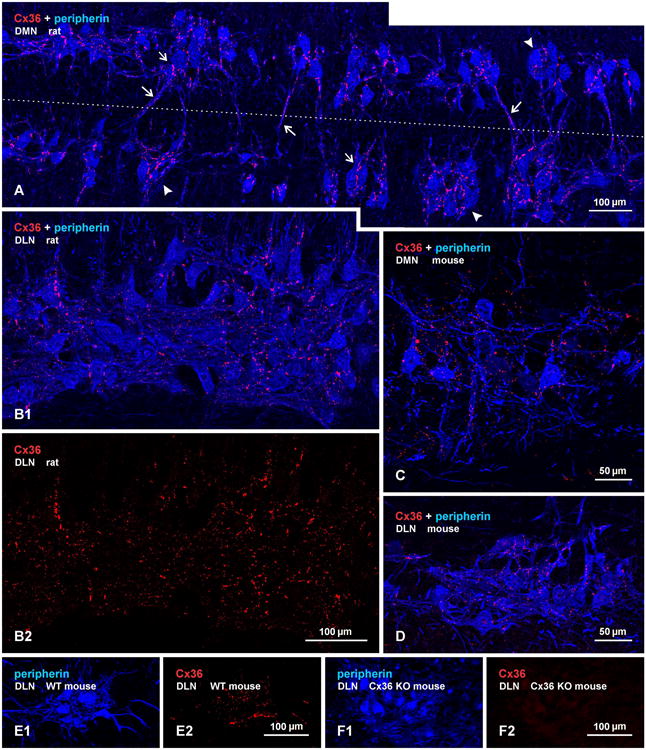
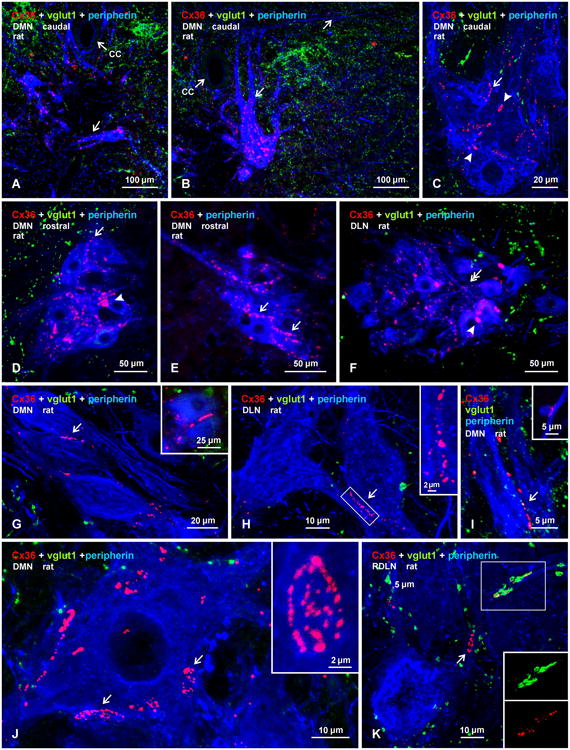
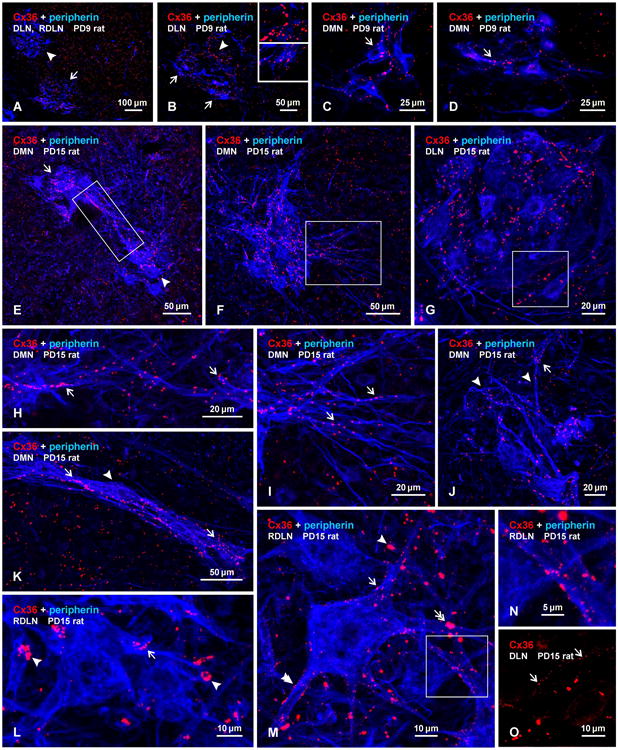
References
-
- Alvarez FJ, Villalba RM, Zerda R, Schneider SP. Vesicular glutamate transporters in the spinal cord, with special reference to sensory primary afferent synapses. J Comp Neurol. 2004;472:257–280. - PubMed
-
- Arasaki K, Kudo N, Nakanishi T. Firing of spinal motoneurons due to electrical interactions in the rat: an in vitro study. Exp Brain Res. 1984;54:437–445. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
