

Eco-evolutionary feedbacks drive species interactions
- PMID: 24304674
- PMCID: PMC3996687
- DOI: 10.1038/ismej.2013.208
Eco-evolutionary feedbacks drive species interactions
Abstract
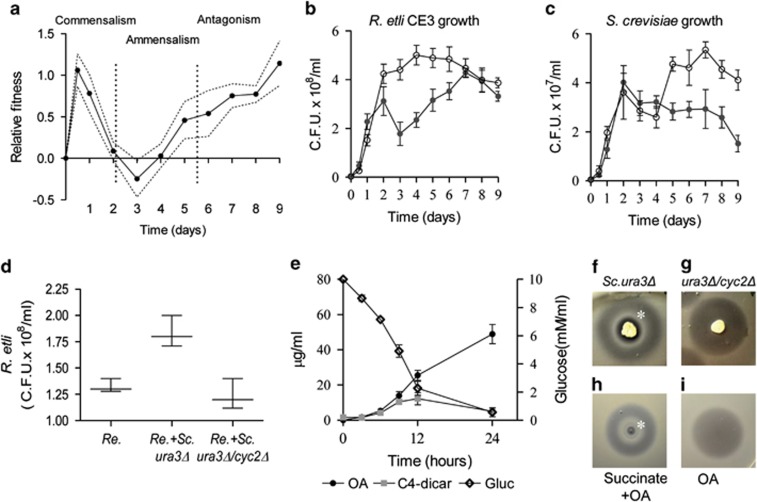
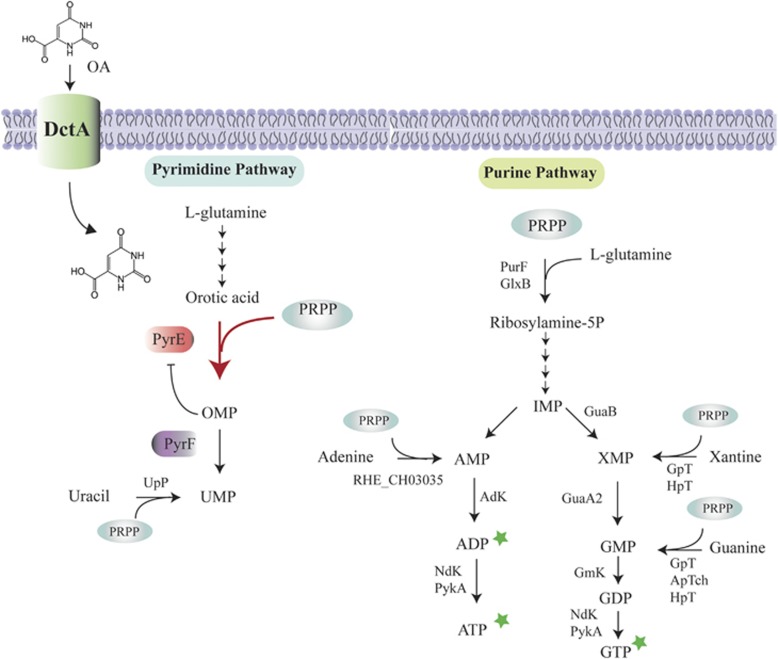
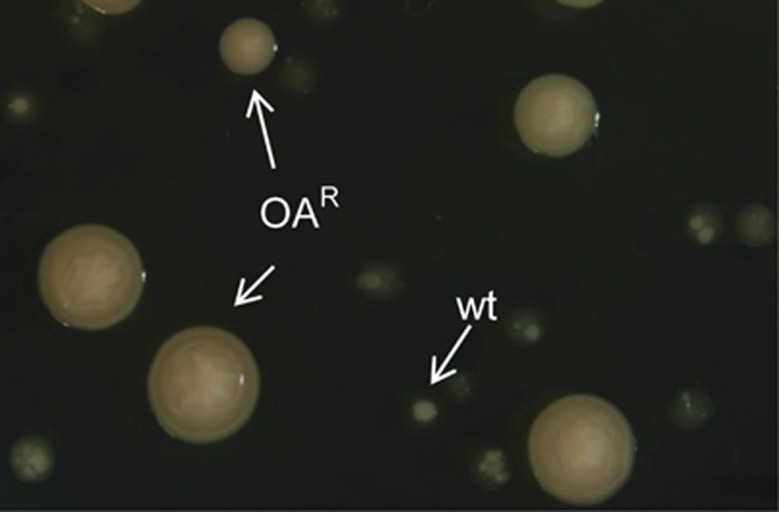
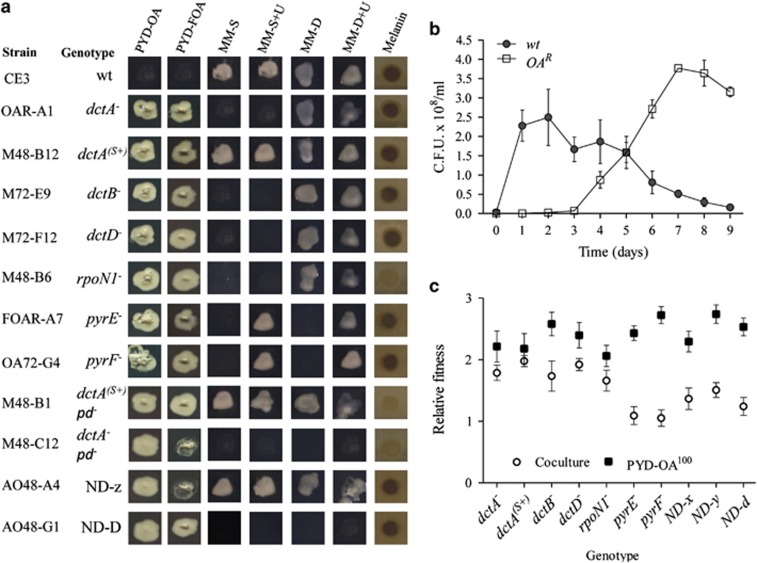
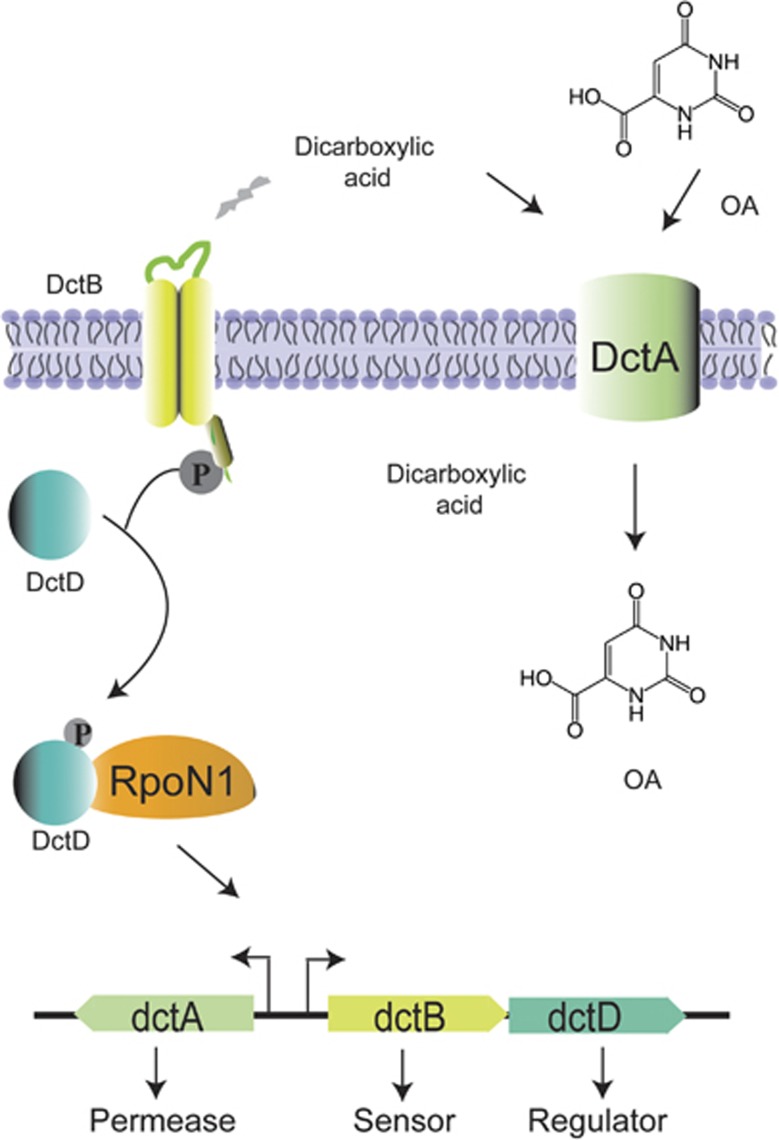
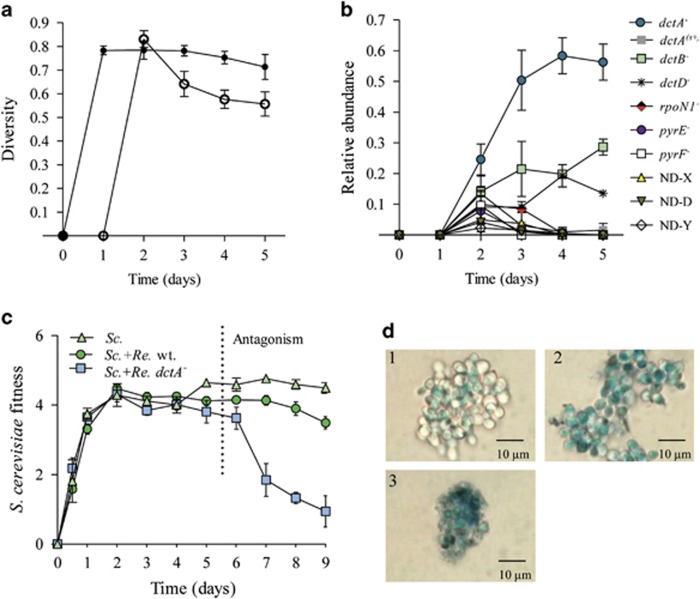
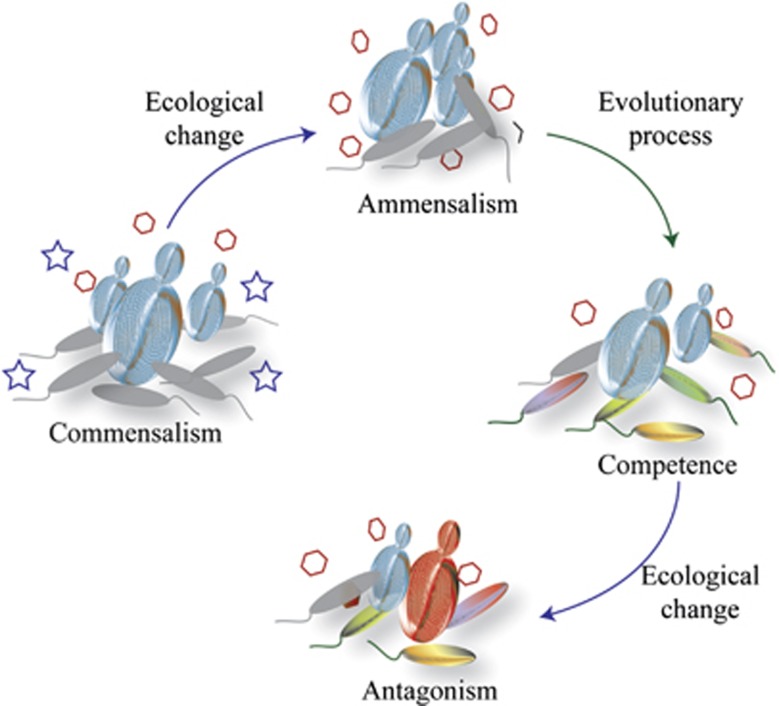
In the biosphere, many species live in close proximity and can thus interact in many different ways. Such interactions are dynamic and fall along a continuum between antagonism and cooperation. Because interspecies interactions are the key to understanding biological communities, it is important to know how species interactions arise and evolve. Here, we show that the feedback between ecological and evolutionary processes has a fundamental role in the emergence and dynamics of species interaction. Using a two-species artificial community, we demonstrate that ecological processes and rapid evolution interact to influence the dynamics of the symbiosis between a eukaryote (Saccharomyces cerevisiae) and a bacterium (Rhizobium etli). The simplicity of our experimental design enables an explicit statement of causality. The niche-constructing activities of the fungus were the key ecological process: it allowed the establishment of a commensal relationship that switched to ammensalism and provided the selective conditions necessary for the adaptive evolution of the bacteria. In this latter state, the bacterial population radiates into more than five genotypes that vary with respect to nutrient transport, metabolic strategies and global regulation. Evolutionary diversification of the bacterial populations has strong effects on the community; the nature of interaction subsequently switches from ammensalism to antagonism where bacteria promote yeast extinction. Our results demonstrate the importance of the evolution-to-ecology pathway in the persistence of interactions and the stability of communities. Thus, eco-evolutionary dynamics have the potential to transform the structure and functioning of ecosystems. Our results suggest that these dynamics should be considered to improve our understanding of beneficial and detrimental host-microbe interactions.
Figures
References
-
- Azam F, Malfatti F. Microbial structuring of marine ecosystems. Nat Rev Microbiol. 2007;5:782–791. - PubMed
-
- Bell G, Gonzalez A. Adaptation and evolutionary rescue in metapopulations experiencing environmental deterioration. Science. 2011;332:1327–1330. - PubMed
-
- Buckling A, Rainey PB. The role of parasites in sympatric and allopatric host diversification. Nature. 2002;420:496–499. - PubMed
Publication types
MeSH terms
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
