

A novel tumor-promoting role for nuclear factor IA in glioblastomas is mediated through negative regulation of p53, p21, and PAI1
- PMID: 24305710
- PMCID: PMC3895381
- DOI: 10.1093/neuonc/not167
A novel tumor-promoting role for nuclear factor IA in glioblastomas is mediated through negative regulation of p53, p21, and PAI1
Abstract
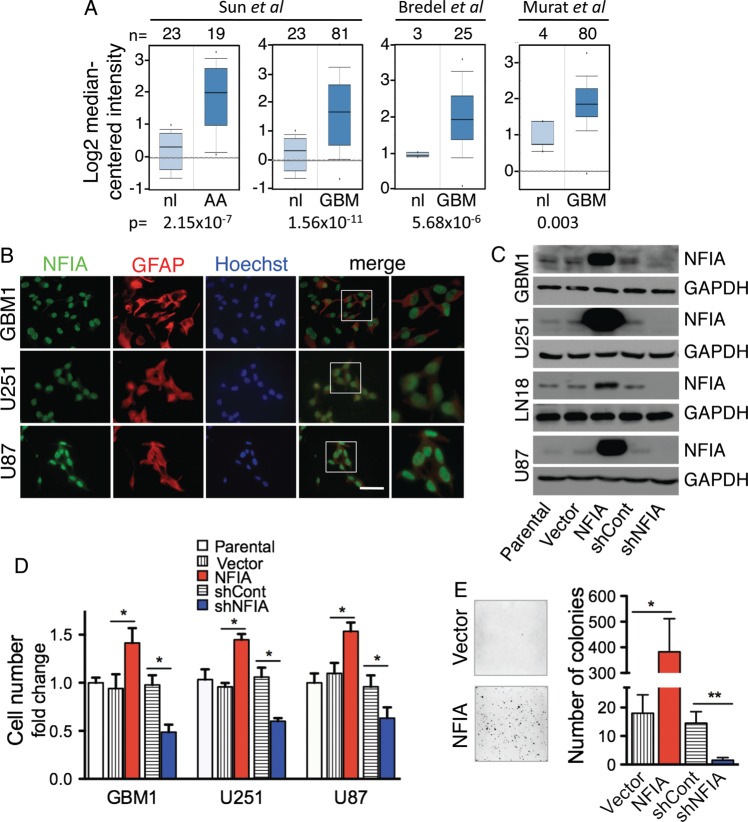
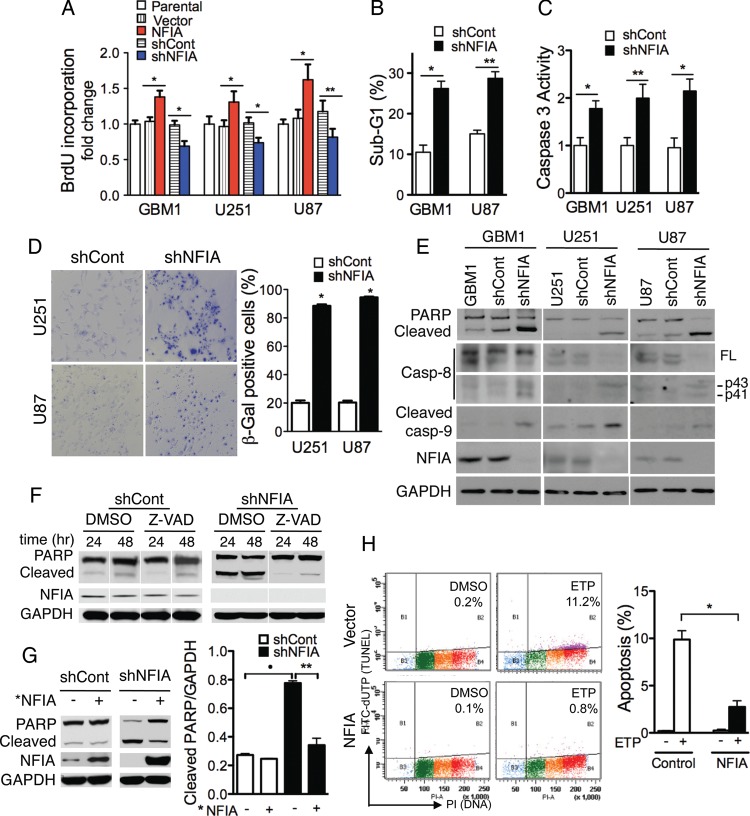
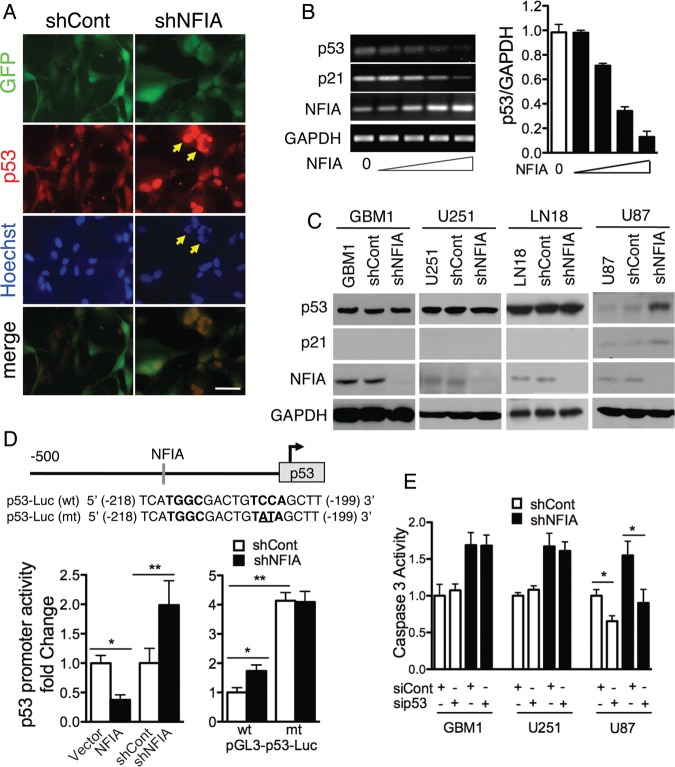
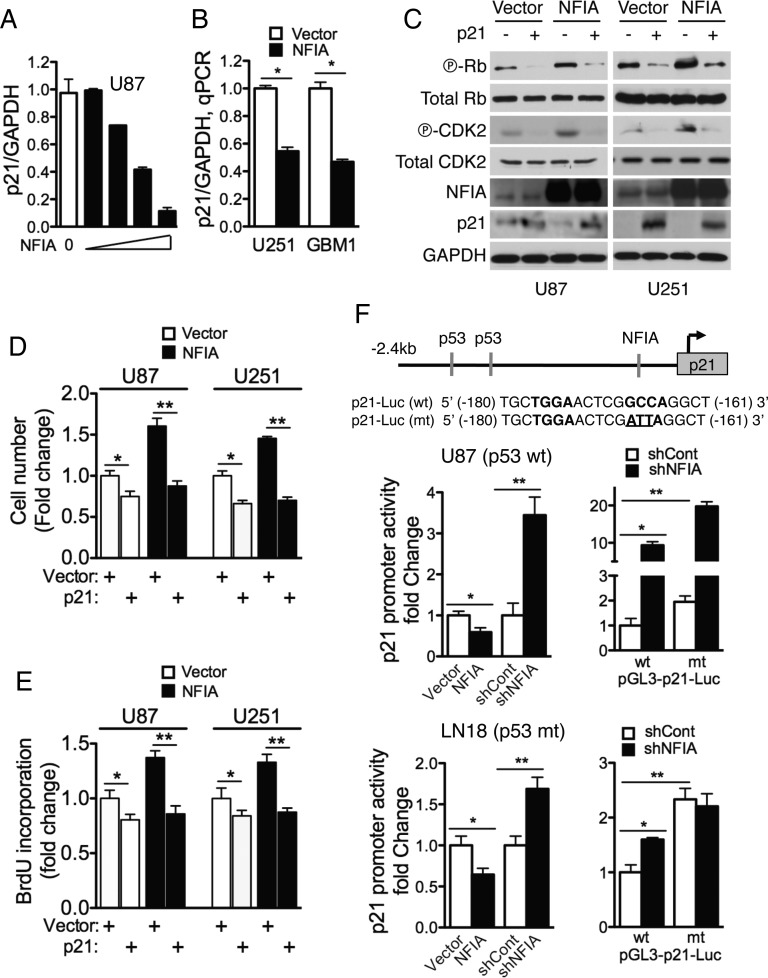
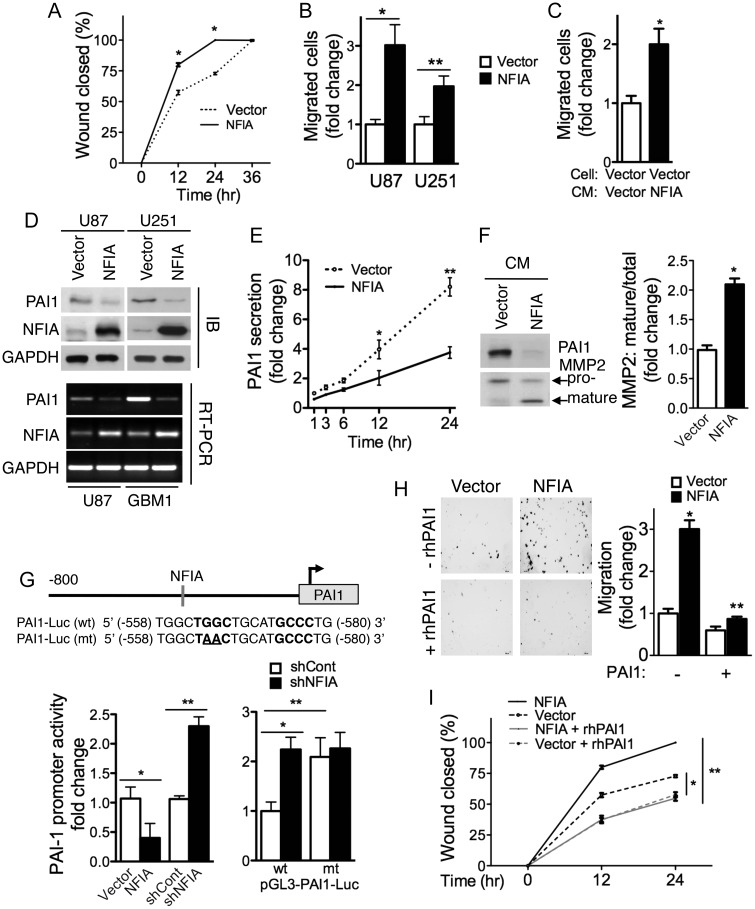
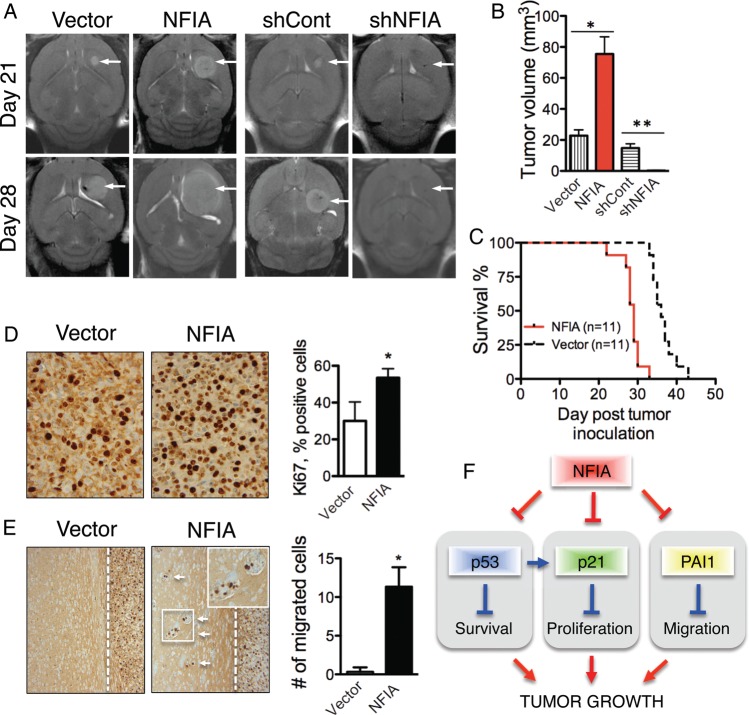
Background Nuclear factor IA (NFIA), a transcription factor and essential regulator in embryonic glial development, is highly expressed in human glioblastoma (GBM) compared with normal brain, but its contribution to GBM and cancer pathogenesis is unknown. Here we demonstrate a novel role for NFIA in promoting growth and migration of GBM and establish the molecular mechanisms mediating these functions. Methods To determine the role of NFIA in glioma, we examined the effects of NFIA in growth, proliferation, apoptosis, and migration. We used gain-of-function (overexpression) and loss-of-function (shRNA knockdown) of NFIA in primary patient-derived GBM cells and established glioma cell lines in culture and in intracranial xenografts in mouse brains. Results Knockdown of native NFIA blocked tumor growth and induced cell death and apoptosis. Complementing this, NFIA overexpression accelerated growth, proliferation, and migration of GBM in cell culture and in mouse brains. These NFIA tumor-promoting effects were mediated via transcriptional repression of p53, p21, and plasminogen activator inhibitor 1 (PAI1) through specific NFIA-recognition sequences in their promoters. Importantly, the effects of NFIA on proliferation and apoptosis were independent of TP53 mutation status, a finding especially relevant for GBM, in which TP53 is frequently mutated. Conclusion NFIA is a modulator of GBM growth and migration, and functions by distinct regulation of critical oncogenic pathways that govern the malignant behavior of GBM.
Keywords: PAI1; glioblastoma (GBM); glioma; nuclear factor IA (NFIA); p21; p53.
Figures
Similar articles
-
Nuclear factor I A promotes temozolomide resistance in glioblastoma via activation of nuclear factor κB pathway.Life Sci. 2019 Nov 1;236:116917. doi: 10.1016/j.lfs.2019.116917. Epub 2019 Oct 12. Life Sci. 2019. PMID: 31614149
-
Repression of Septin9 and Septin2 suppresses tumor growth of human glioblastoma cells.Cell Death Dis. 2018 May 1;9(5):514. doi: 10.1038/s41419-018-0547-4. Cell Death Dis. 2018. PMID: 29724999 Free PMC article.
-
A novel NFIA-NFκB feed-forward loop contributes to glioblastoma cell survival.Neuro Oncol. 2017 Apr 1;19(4):524-534. doi: 10.1093/neuonc/now233. Neuro Oncol. 2017. PMID: 27994064 Free PMC article.
-
The p53 Pathway in Glioblastoma.Cancers (Basel). 2018 Sep 1;10(9):297. doi: 10.3390/cancers10090297. Cancers (Basel). 2018. PMID: 30200436 Free PMC article. Review.
-
Sex-Specific Differences in Glioblastoma.Cells. 2021 Jul 14;10(7):1783. doi: 10.3390/cells10071783. Cells. 2021. PMID: 34359952 Free PMC article. Review.
Cited by
-
A Glance of p53 Functions in Brain Development, Neural Stem Cells, and Brain Cancer.Biology (Basel). 2020 Sep 11;9(9):285. doi: 10.3390/biology9090285. Biology (Basel). 2020. PMID: 32932978 Free PMC article. Review.
-
Mechanisms of Cell Cycle Arrest and Apoptosis in Glioblastoma.Biomedicines. 2022 Feb 28;10(3):564. doi: 10.3390/biomedicines10030564. Biomedicines. 2022. PMID: 35327366 Free PMC article. Review.
-
An MDM2 inhibitor achieves synergistic cytotoxic effects with adenoviruses lacking E1B55kDa gene on mesothelioma with the wild-type p53 through augmenting NFI expression.Cell Death Dis. 2021 Jul 2;12(7):663. doi: 10.1038/s41419-021-03934-y. Cell Death Dis. 2021. PMID: 34230456 Free PMC article.
-
Disruptive chemicals, senescence and immortality.Carcinogenesis. 2015 Jun;36 Suppl 1(Suppl 1):S19-37. doi: 10.1093/carcin/bgv029. Carcinogenesis. 2015. PMID: 26106138 Free PMC article. Review.
-
Nuclear Factor I Represses the Notch Effector HEY1 in Glioblastoma.Neoplasia. 2018 Oct;20(10):1023-1037. doi: 10.1016/j.neo.2018.08.007. Epub 2018 Sep 6. Neoplasia. 2018. PMID: 30195713 Free PMC article.
References
-
- Stupp R, Hegi ME, Mason WP, et al. Effects of radiotherapy with concomitant and adjuvant temozolomide versus radiotherapy alone on survival in glioblastoma in a randomised phase III study: 5-year analysis of the EORTC-NCIC trial. Lancet Oncol. 2009;10(5):459–466. - PubMed
-
- Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646–674. - PubMed
-
- Siebzehnrubl FA, Reynolds BA, Vescovi A, Steindler DA, Deleyrolle LP. The origins of glioma: E Pluribus Unum? Glia. 2011;59(8):1135–1147. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
