

Drosophila intermediate neural progenitors produce lineage-dependent related series of diverse neurons
- PMID: 24306106
- PMCID: PMC3879808
- DOI: 10.1242/dev.103069
Drosophila intermediate neural progenitors produce lineage-dependent related series of diverse neurons
Erratum in
- Development. 2014 Jan;141(2):488
Abstract
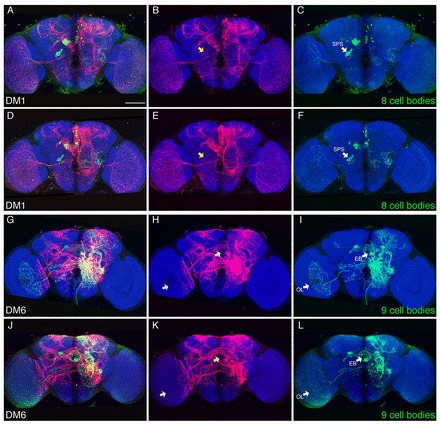
Drosophila type II neuroblasts (NBs), like mammalian neural stem cells, deposit neurons through intermediate neural progenitors (INPs) that can each produce a series of neurons. Both type II NBs and INPs exhibit age-dependent expression of various transcription factors, potentially specifying an array of diverse neurons by combinatorial temporal patterning. Not knowing which mature neurons are made by specific INPs, however, conceals the actual variety of neuron types and limits further molecular studies. Here we mapped neurons derived from specific type II NB lineages and found that sibling INPs produced a morphologically similar but temporally regulated series of distinct neuron types. This suggests a common fate diversification program operating within each INP that is modulated by NB age to generate slightly different sets of diverse neurons based on the INP birth order. Analogous mechanisms might underlie the expansion of neuron diversity via INPs in mammalian brain.
Keywords: Adult brain; Cell lineage analysis; Drosophila type II neuroblasts; Intermediate neural progenitors; Neuronal cell fate; Temporal identity.
Figures



References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
