

The FupA/B protein uniquely facilitates transport of ferrous iron and siderophore-associated ferric iron across the outer membrane of Francisella tularensis live vaccine strain
- PMID: 24307666
- PMCID: PMC3919536
- DOI: 10.1099/mic.0.072835-0
The FupA/B protein uniquely facilitates transport of ferrous iron and siderophore-associated ferric iron across the outer membrane of Francisella tularensis live vaccine strain
Abstract
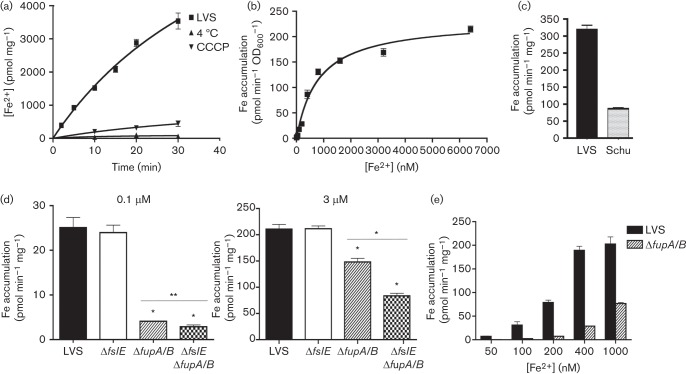
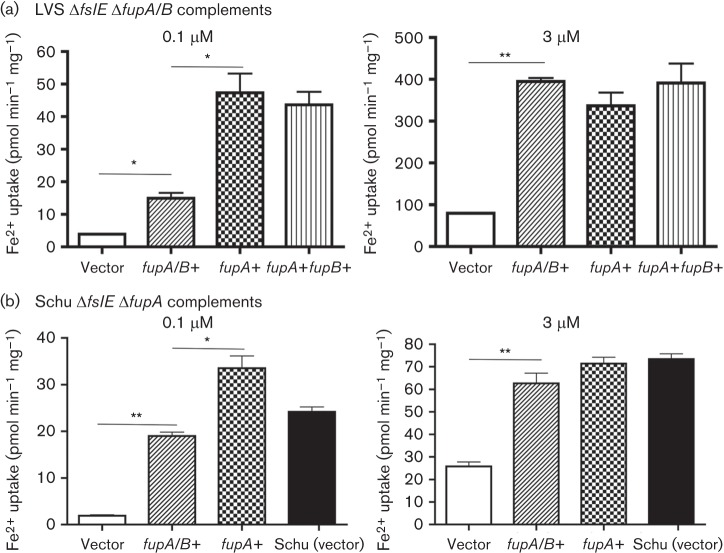
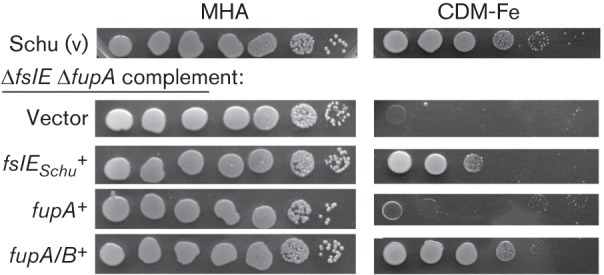
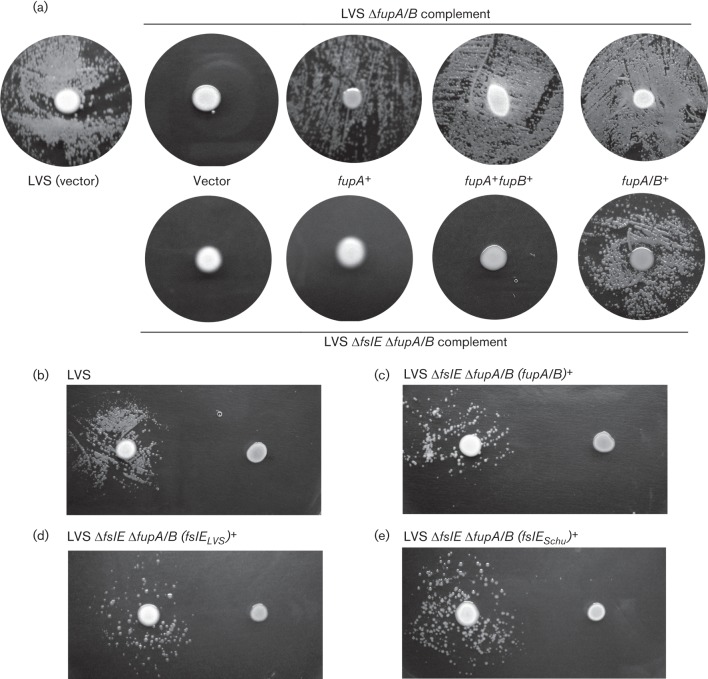
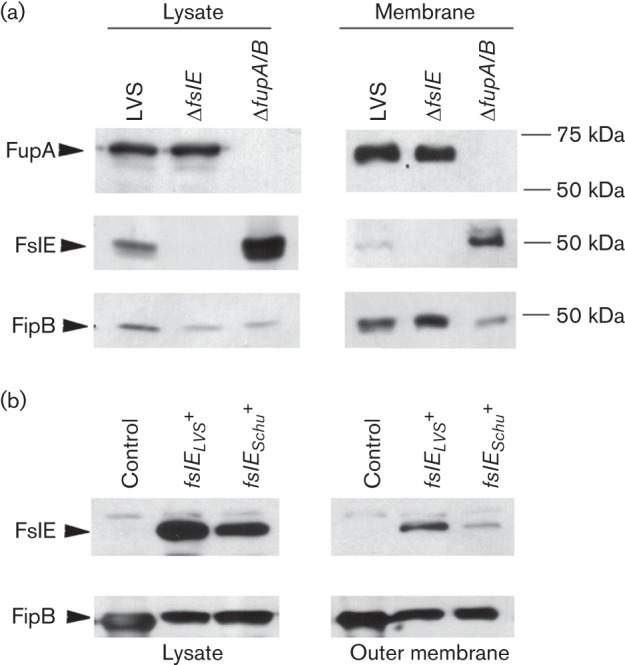
Francisella tularensis is a highly infectious Gram-negative pathogen that replicates intracellularly within the mammalian host. One of the factors associated with virulence of F. tularensis is the protein FupA that mediates high-affinity transport of ferrous iron across the outer membrane. Together with its paralogue FslE, a siderophore-ferric iron transporter, FupA supports survival of the pathogen in the host by providing access to the essential nutrient iron. The FupA orthologue in the attenuated live vaccine strain (LVS) is encoded by the hybrid gene fupA/B, the product of an intergenic recombination event that significantly contributes to attenuation of the strain. We used (55)Fe transport assays with mutant strains complemented with the different paralogues to show that the FupA/B protein of LVS retains the capacity for high-affinity transport of ferrous iron, albeit less efficiently than FupA of virulent strain Schu S4. (55)Fe transport assays using purified siderophore and siderophore-dependent growth assays on iron-limiting agar confirmed previous findings that FupA/B also contributes to siderophore-mediated ferric iron uptake. These assays further demonstrated that the LVS FslE protein is a weaker siderophore-ferric iron transporter than the orthologue from Schu S4, and may be a result of the sequence variation between the two proteins. Our results indicate that iron-uptake mechanisms in LVS differ from those in Schu S4 and that functional differences in the outer membrane iron transporters have distinct effects on growth under iron limitation.
Figures
References
-
- Broekhuijsen M., Larsson P., Johansson A., Byström M., Eriksson U., Larsson E., Prior R. G., Sjöstedt A., Titball R. W., Forsman M. (2003). Genome-wide DNA microarray analysis of Francisella tularensis strains demonstrates extensive genetic conservation within the species but identifies regions that are unique to the highly virulent F. tularensis subsp. tularensis. J Clin Microbiol 41, 2924–2931. 10.1128/JCM.41.7.2924-2931.2003 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical