

Gamma band activity in the RAS-intracellular mechanisms
- PMID: 24309750
- PMCID: PMC4013218
- DOI: 10.1007/s00221-013-3794-8
Gamma band activity in the RAS-intracellular mechanisms
Erratum in
- Exp Brain Res. 2014 May;232(5):1523
Abstract
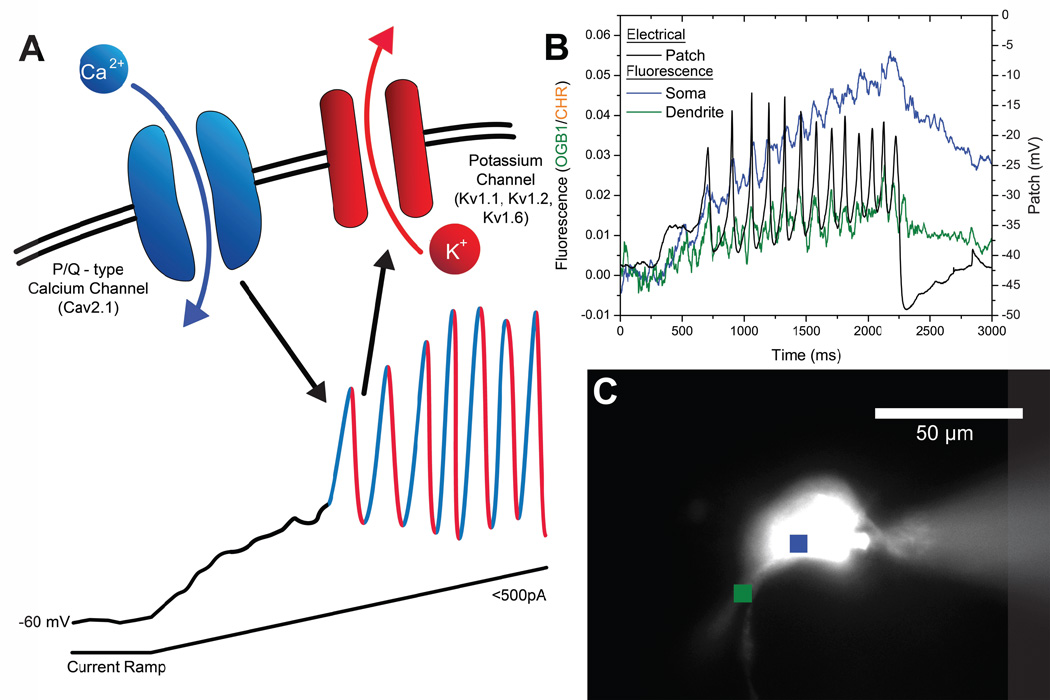
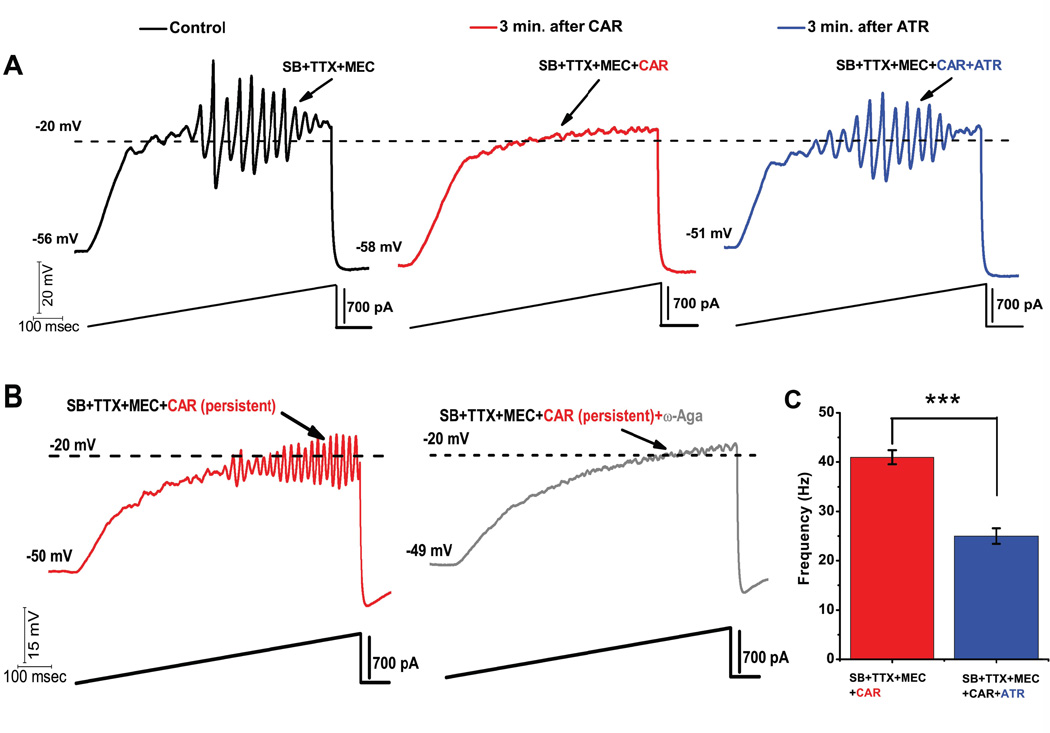
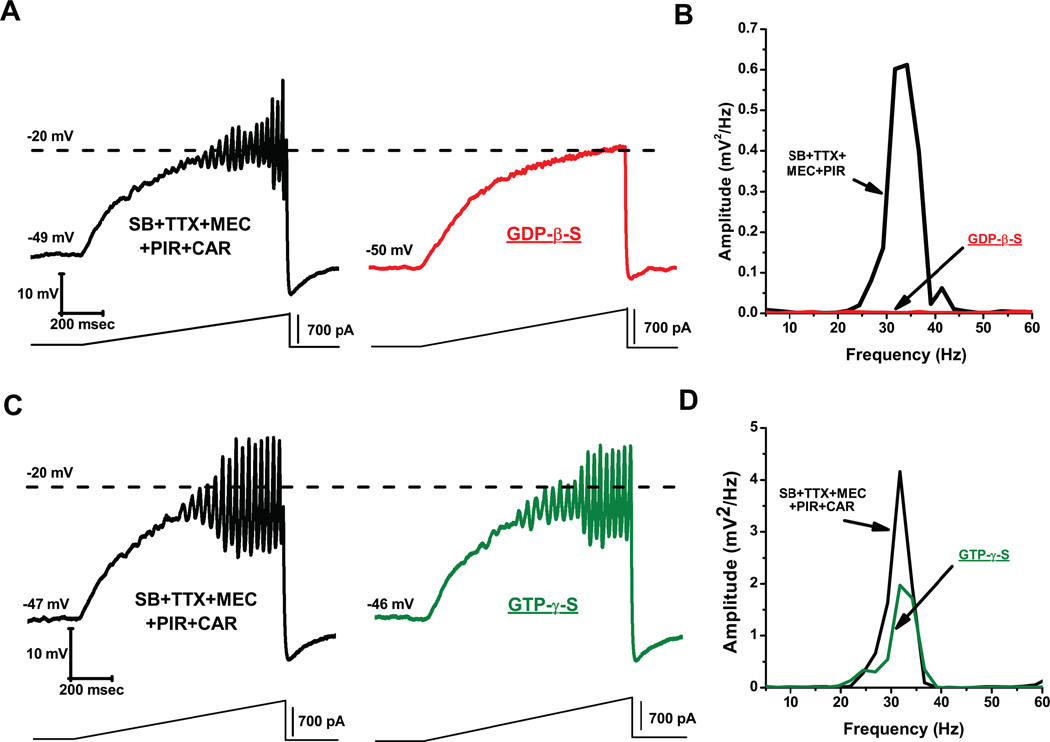
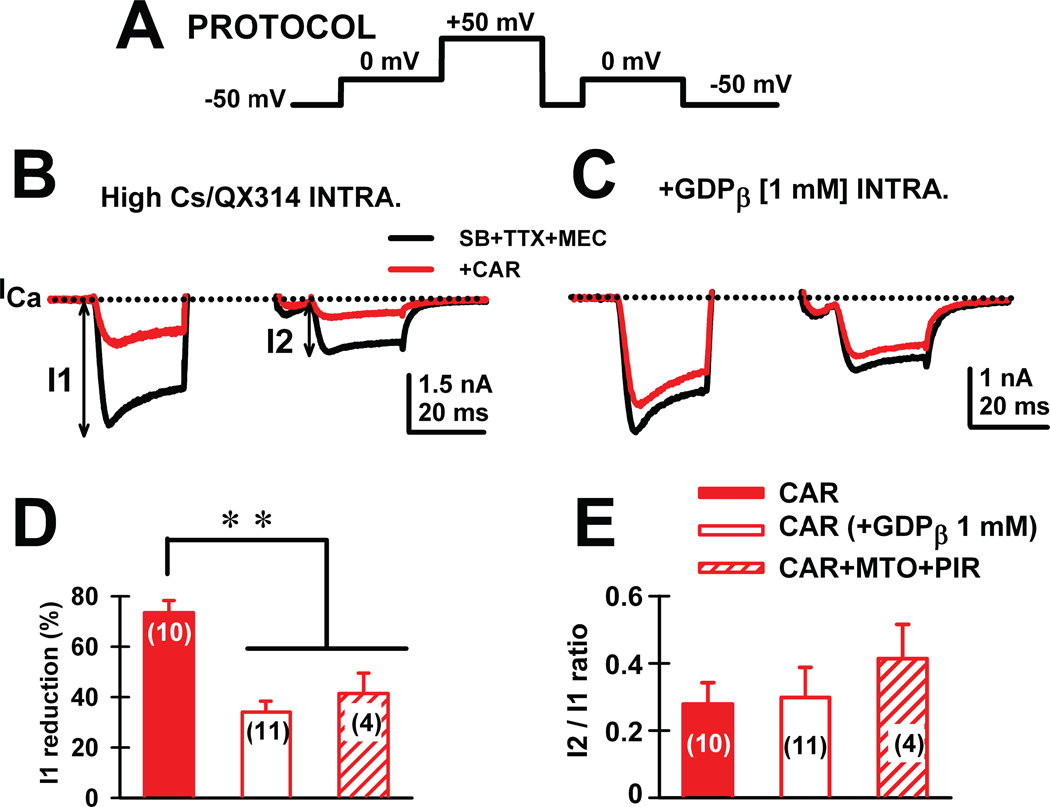
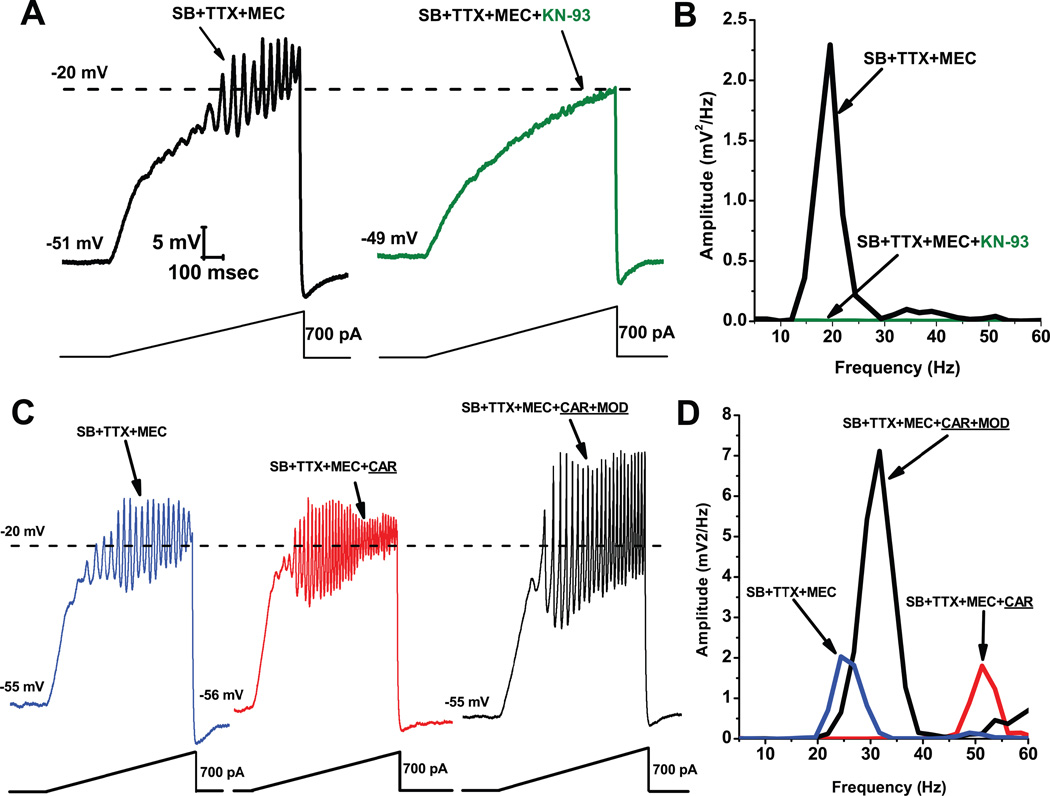
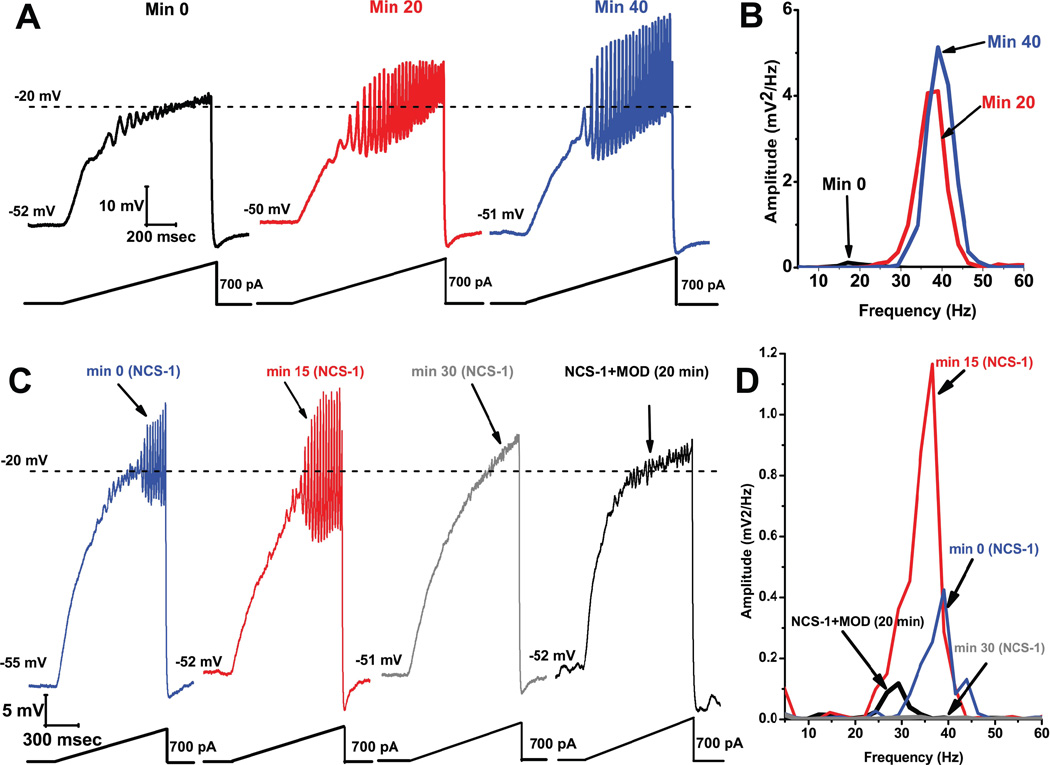
Gamma band activity participates in sensory perception, problem solving, and memory. This review considers recent evidence showing that cells in the reticular activating system (RAS) exhibit gamma band activity, and describes the intrinsic membrane properties behind such manifestation. Specifically, we discuss how cells in the mesopontine pedunculopontine nucleus, intralaminar parafascicular nucleus, and pontine SubCoeruleus nucleus dorsalis all fire in the gamma band range when maximally activated, but no higher. The mechanisms involve high-threshold, voltage-dependent P/Q-type calcium channels, or sodium-dependent subthreshold oscillations. Rather than participating in the temporal binding of sensory events as in the cortex, gamma band activity in the RAS may participate in the processes of preconscious awareness and provide the essential stream of information for the formulation of many of our actions. We address three necessary next steps resulting from these discoveries: an intracellular mechanism responsible for maintaining gamma band activity based on persistent G-protein activation, separate intracellular pathways that differentiate between gamma band activity during waking versus during REM sleep, and an intracellular mechanism responsible for the dysregulation in gamma band activity in schizophrenia. These findings open several promising research avenues that have not been thoroughly explored. What are the effects of sleep or REM sleep deprivation on these RAS mechanisms? Are these mechanisms involved in memory processing during waking and/or during REM sleep? Does gamma band processing differ during waking versus REM sleep after sleep or REM sleep deprivation?
Figures
References
-
- Aserinsky E, Kleitman N. Regularly occurring periods of eye motility, and concomitant phenomena, during sleep. Science. 1953;118:273–274. - PubMed
-
- Baghdoyan HA, Rodrigo-Angulo ML, McCarley RW, Hobson JA. A neuroanatomical gradient in the pontine tegmentum for the cholinoceptive induction of desynchronized sleep signs. Brain Res. 1987;414:245–261. - PubMed
-
- Berger M, Riemann D. Symposium: Normal and abnormal REM sleep regulation: REM sleep in depression- an overview. J Sleep Res. 1993;2:211–223. - PubMed
-
- Bergson C, Levenson R, Goldman-Rakic P, Lidow MS. Dopamine receptor-interacting proteins: the Ca2+ connection in dopamine signaling. Trends Pharmacol Sci. 2003;24:486–492. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
