

Extracellular vesicles modulate the glioblastoma microenvironment via a tumor suppression signaling network directed by miR-1
- PMID: 24310399
- PMCID: PMC3928601
- DOI: 10.1158/0008-5472.CAN-13-2650
Extracellular vesicles modulate the glioblastoma microenvironment via a tumor suppression signaling network directed by miR-1
Abstract
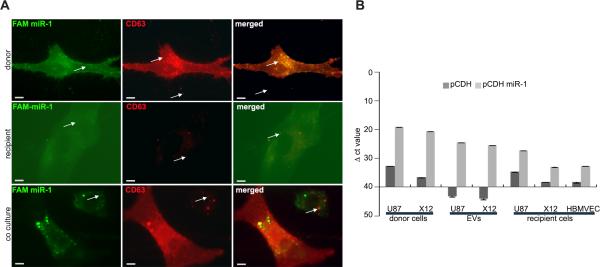
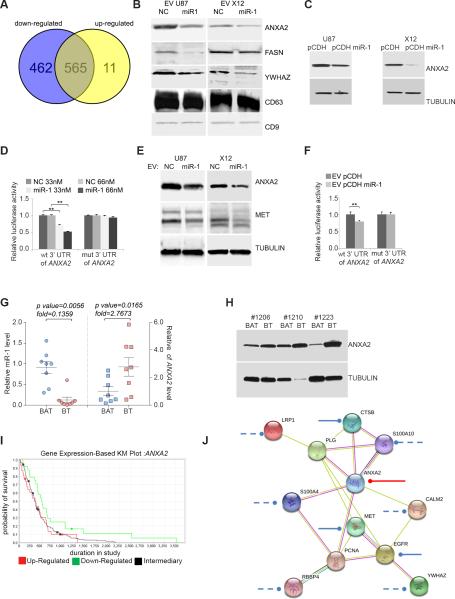
Extracellular vesicles have emerged as important mediators of intercellular communication in cancer, including by conveying tumor-promoting microRNAs between cells, but their regulation is poorly understood. In this study, we report the findings of a comparative microRNA profiling and functional analysis in human glioblastoma that identifies miR-1 as an orchestrator of extracellular vesicle function and glioblastoma growth and invasion. Ectopic expression of miR-1 in glioblastoma cells blocked in vivo growth, neovascularization, and invasiveness. These effects were associated with a role for miR-1 in intercellular communication in the microenvironment mediated by extracellular vesicles released by cancer stem-like glioblastoma cells. An extracellular vesicle-dependent phenotype defined by glioblastoma invasion, neurosphere growth, and endothelial tube formation was mitigated by loading miR-1 into glioblastoma-derived extracellular vesicles. Protein cargo in extracellular vesicles was characterized to learn how miR-1 directed extracellular vesicle function. The mRNA encoding Annexin A2 (ANXA2), one of the most abundant proteins in glioblastoma-derived extracellular vesicles, was found to be a direct target of miR-1 control. In addition, extracellular vesicle-derived miR-1 along with other ANXA2 extracellular vesicle networking partners targeted multiple pro-oncogenic signals in cells within the glioblastoma microenvironment. Together, our results showed how extracellular vesicle signaling promotes the malignant character of glioblastoma and how ectopic expression of miR-1 can mitigate this character, with possible implications for how to develop a unique miRNA-based therapy for glioblastoma management.
Figures
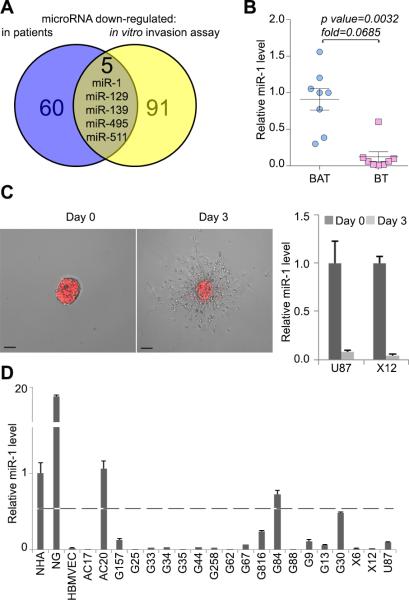
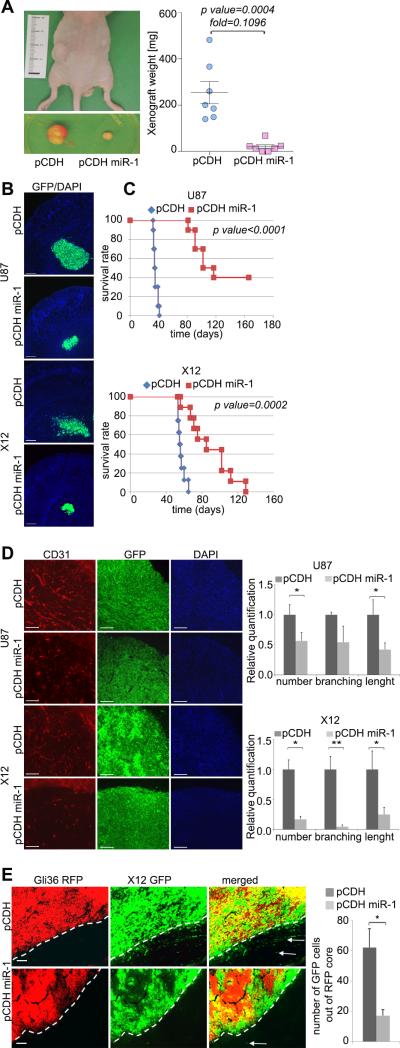
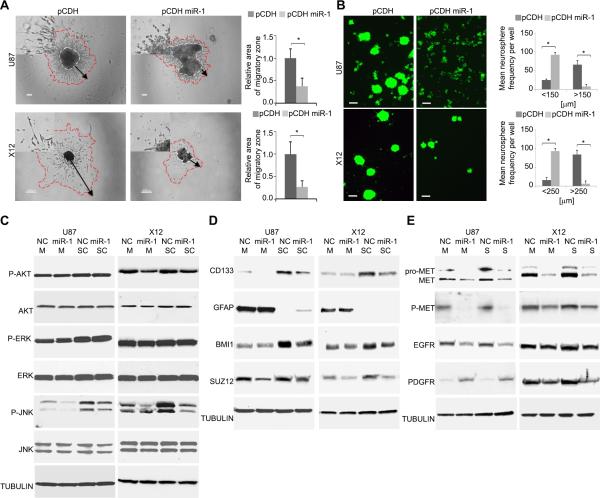
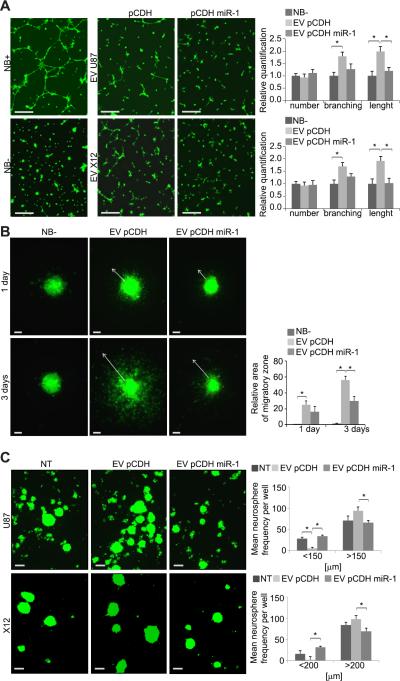
References
-
- Hess KR, Broglio KR, Bondy ML. Adult glioma incidence trends in the United States, 1977-2000. Cancer. 2004;101:2293–9. - PubMed
-
- Stupp R, Mason WP, van den Bent MJ, Weller M, Fisher B, Taphoorn MJ, et al. Radiotherapy plus concomitant and adjuvant temozolomide for glioblastoma. N Engl J Med. 2005;352:987–96. - PubMed
-
- Furnari FB, Fenton T, Bachoo RM, Mukasa A, Stommel JM, Stegh A, et al. Malignant astrocytic glioma: genetics, biology, and paths to treatment. Genes Dev. 2007;21:2683–710. - PubMed
-
- Lefranc F, Brotchi J, Kiss R. Possible future issues in the treatment of glioblastomas: special emphasis on cell migration and the resistance of migrating glioblastoma cells to apoptosis. J Clin Oncol. 2005;23:2411–22. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
