

miR-9: a versatile regulator of neurogenesis
- PMID: 24312010
- PMCID: PMC3834235
- DOI: 10.3389/fncel.2013.00220
miR-9: a versatile regulator of neurogenesis
Abstract
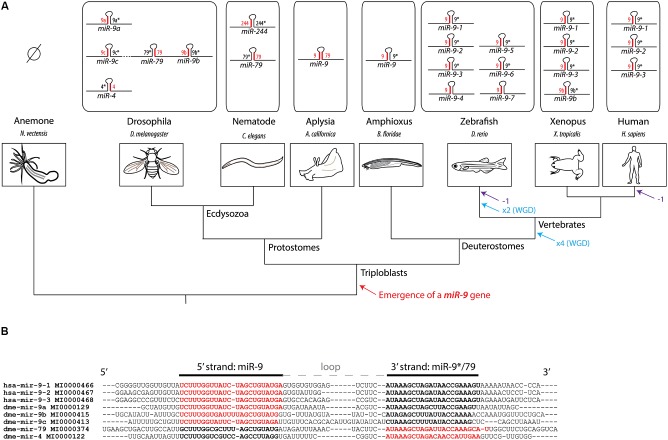
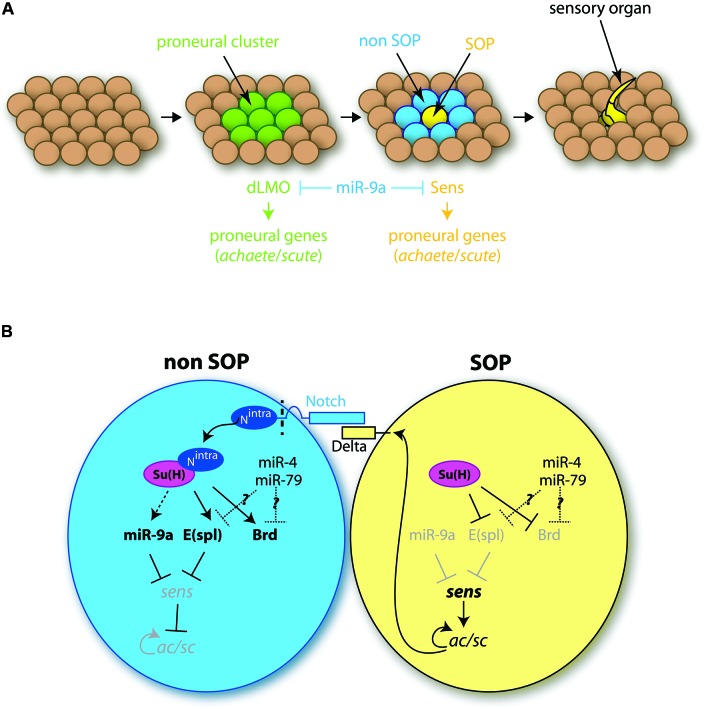
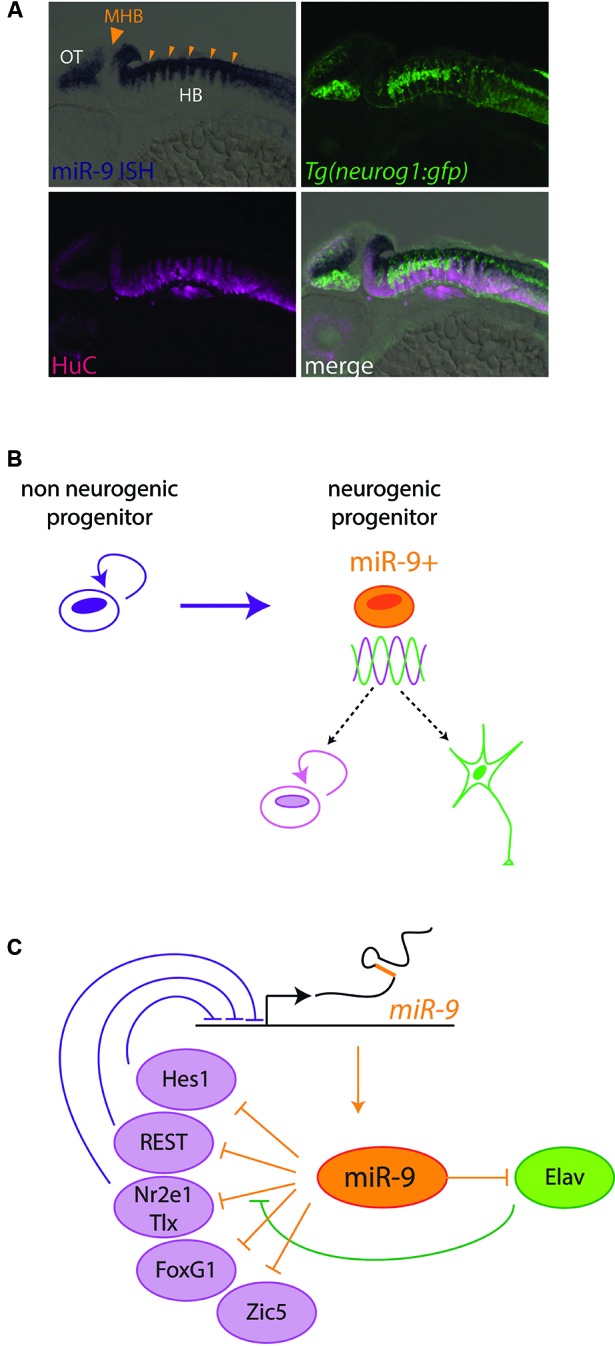
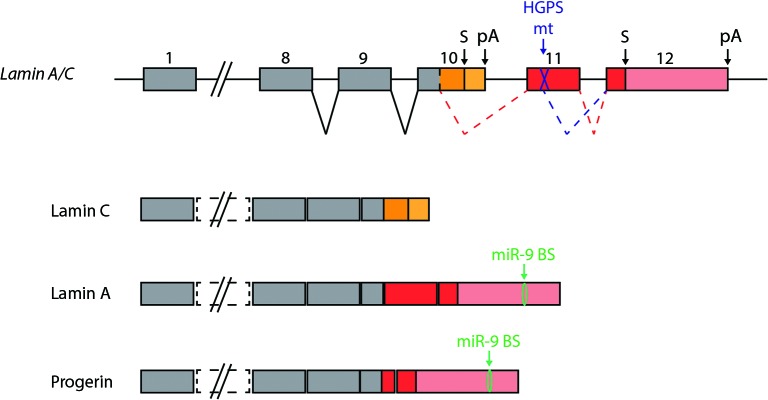
Soon after its discovery, microRNA-9 (miR-9) attracted the attention of neurobiologists, since it is one of the most highly expressed microRNAs in the developing and adult vertebrate brain. Functional analyses in different vertebrate species have revealed a prominent role of this microRNA in balancing proliferation in embryonic neural progenitor populations. Key transcriptional regulators such as FoxG1, Hes1 or Tlx, were identified as direct targets of miR-9, placing it at the core of the gene network controlling the progenitor state. Recent data also suggest that this function could extend to adult neural stem cells. Other studies point to a role of miR-9 in differentiated neurons. Moreover miR-9 has been implicated in human brain pathologies, either displaying a protective role, such as in Progeria, or participating in disease progression in brain cancers. Altogether functional studies highlight a prominent feature of this highly conserved microRNA, its functional versatility, both along its evolutionary history and across cellular contexts.
Keywords: embryonic progenitors; microRNA-9; neural stem cells; neurogenesis; proliferation.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases