

Is the fluid mosaic (and the accompanying raft hypothesis) a suitable model to describe fundamental features of biological membranes? What may be missing?
- PMID: 24312108
- PMCID: PMC3826152
- DOI: 10.3389/fpls.2013.00457
Is the fluid mosaic (and the accompanying raft hypothesis) a suitable model to describe fundamental features of biological membranes? What may be missing?
Abstract
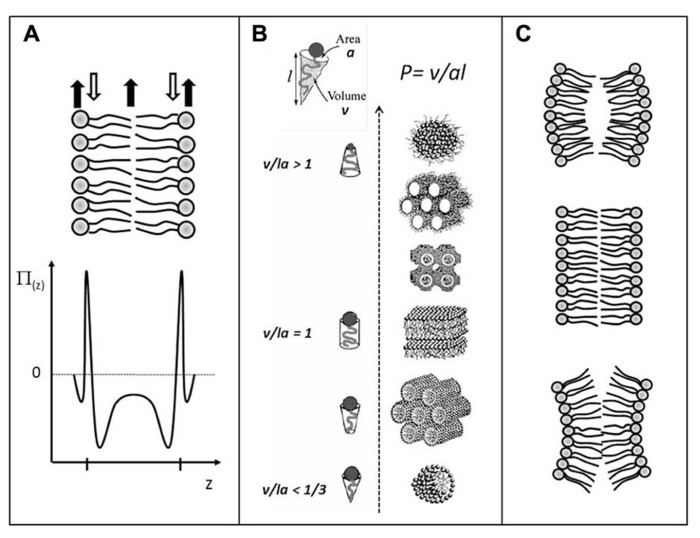
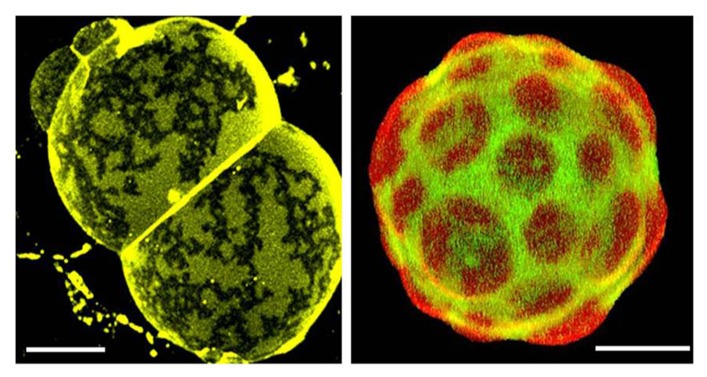
The structure, dynamics, and stability of lipid bilayers are controlled by thermodynamic forces, leading to overall tensionless membranes with a distinct lateral organization and a conspicuous lateral pressure profile. Bilayers are also subject to built-in curvature-stress instabilities that may be released locally or globally in terms of morphological changes leading to the formation of non-lamellar and curved structures. A key controller of the bilayer's propensity to form curved structures is the average molecular shape of the different lipid molecules. Via the curvature stress, molecular shape mediates a coupling to membrane-protein function and provides a set of physical mechanisms for formation of lipid domains and laterally differentiated regions in the plane of the membrane. Unfortunately, these relevant physical features of membranes are often ignored in the most popular models for biological membranes. Results from a number of experimental and theoretical studies emphasize the significance of these fundamental physical properties and call for a refinement of the fluid mosaic model (and the accompanying raft hypothesis).
Keywords: fluid mosaic model; membrane compositional fluctuations; membrane curvature; membrane domains; membrane lateral organization; membrane lateral pressure profile; raft hypothesis.
Figures
References
-
- Bagatolli L. A. (2012). “5.3 Membrane domains and their relevance to the organization of biological membranes,” in Comprehensive Biophysics ed. Edward H. E. (Amsterdam: Elsevier; ) 16–36
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous