

Mutational analysis of sclerostin shows importance of the flexible loop and the cystine-knot for Wnt-signaling inhibition
- PMID: 24312339
- PMCID: PMC3843708
- DOI: 10.1371/journal.pone.0081710
Mutational analysis of sclerostin shows importance of the flexible loop and the cystine-knot for Wnt-signaling inhibition
Erratum in
- PLoS One. 2014;9(1). doi:10.1371/annotation/067f1197-3290-4d1e-8f77-4d3996a6b9e8
Abstract
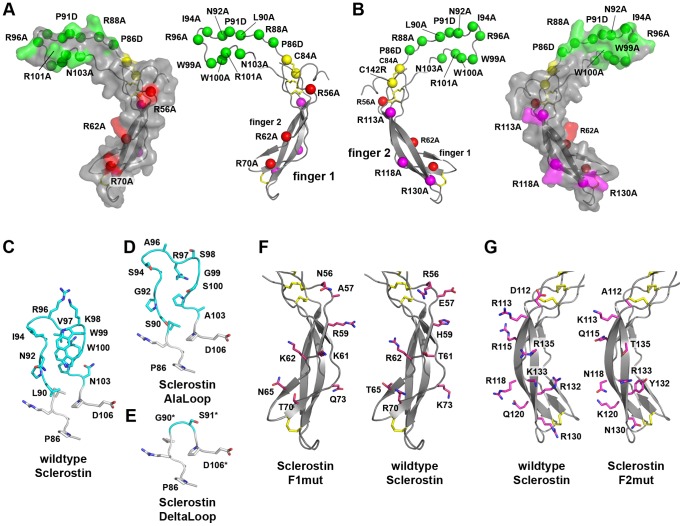
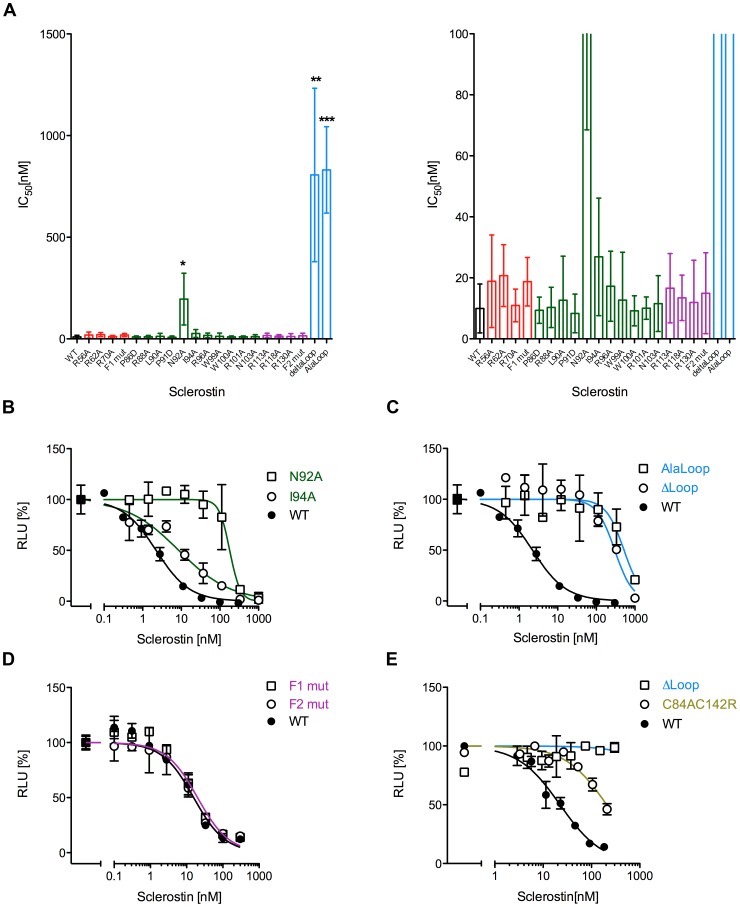
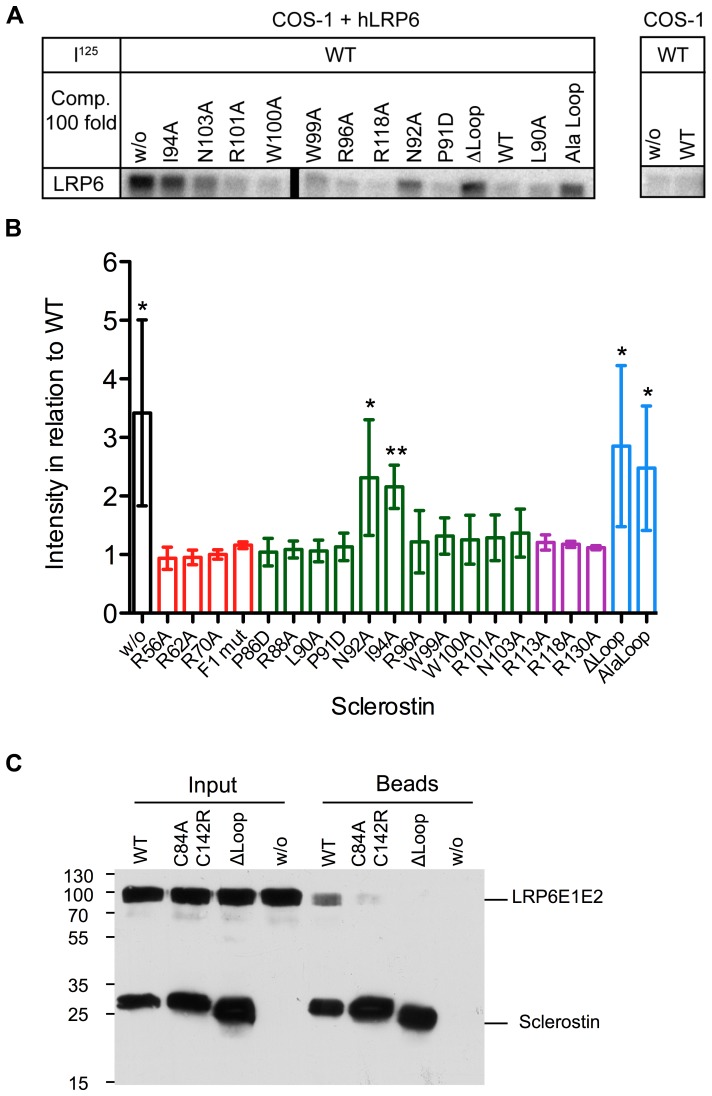
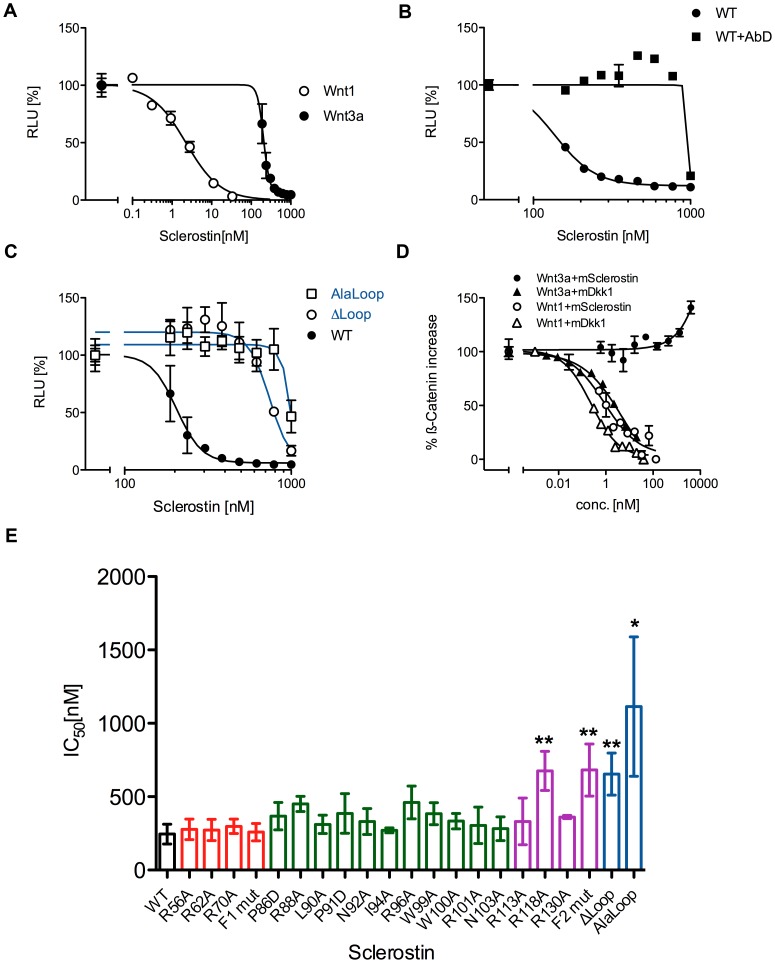
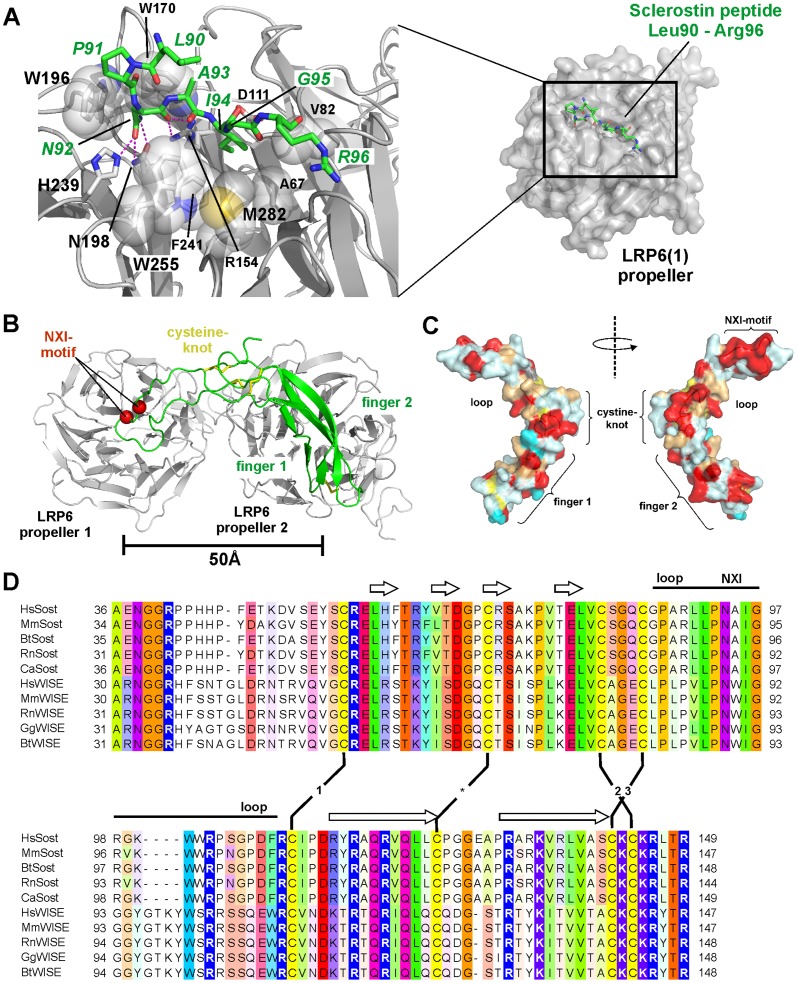
The cystine-knot containing protein Sclerostin is an important negative regulator of bone growth and therefore represents a promising therapeutic target. It exerts its biological task by inhibiting the Wnt (wingless and int1) signaling pathway, which participates in bone formation by promoting the differentiation of mesenchymal stem cells to osteoblasts. The core structure of Sclerostin consists of three loops with the first and third loop (Finger 1 and Finger 2) forming a structured β-sheet and the second loop being unstructured and highly flexible. Biochemical data showed that the flexible loop is important for binding of Sclerostin to Wnt co-receptors of the low-density lipoprotein related-protein family (LRP), by interacting with the Wnt co-receptors LRP5 or -6 it inhibits Wnt signaling. To further examine the structural requirements for Wnt inhibition, we performed an extensive mutational study within all three loops of the Sclerostin core domain involving single and multiple mutations as well as truncation of important regions. By this approach we could confirm the importance of the second loop and especially of amino acids Asn92 and Ile94 for binding to LRP6. Based on a Sclerostin variant found in a Turkish family suffering from Sclerosteosis we generated a Sclerostin mutant with cysteines 84 and 142 exchanged thereby removing the third disulfide bond of the cystine-knot. This mutant binds to LRP6 with reduced binding affinity and also exhibits a strongly reduced inhibitory activity against Wnt1 thereby showing that also elements outside the flexible loop are important for inhibition of Wnt by Sclerostin. Additionally, we examined the effect of the mutations on the inhibition of two different Wnt proteins, Wnt3a and Wnt1. We could detect clear differences in the inhibition of these proteins, suggesting that the mechanism by which Sclerostin antagonizes Wnt1 and Wnt3a is fundamentally different.
Conflict of interest statement
Figures
References
-
- Raisz LG (1999) Physiology and pathophysiology of bone remodeling. Clinical chemistry 45: 1353–1358. - PubMed
-
- Bonewald LF (2007) Osteocytes as Dynamic Multifunctional Cells. Annals of the New York Academy of Sciences 1116: 281–290. - PubMed
-
- Teitelbaum SL (2000) Bone Resorption by Osteoclasts. Science 289: 1504–1508. - PubMed
-
- Katagiri T, Takahashi N (2002) Regulatory mechanisms of osteoblast and osteoclast differentiation. Oral Diseases 8: 147–159. - PubMed
-
- Day TF, Guo X, Garrett-Beal L, Yang Y (2005) Wnt/β-Catenin Signaling in Mesenchymal Progenitors Controls Osteoblast and Chondrocyte Differentiation during Vertebrate Skeletogenesis. Developmental Cell 8: 739–750. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
