

Neuropilar projections of the anterior gastric receptor neuron in the stomatogastric ganglion of the Jonah crab, Cancer borealis
- PMID: 24312448
- PMCID: PMC3848923
- DOI: 10.1371/journal.pone.0079306
Neuropilar projections of the anterior gastric receptor neuron in the stomatogastric ganglion of the Jonah crab, Cancer borealis
Abstract
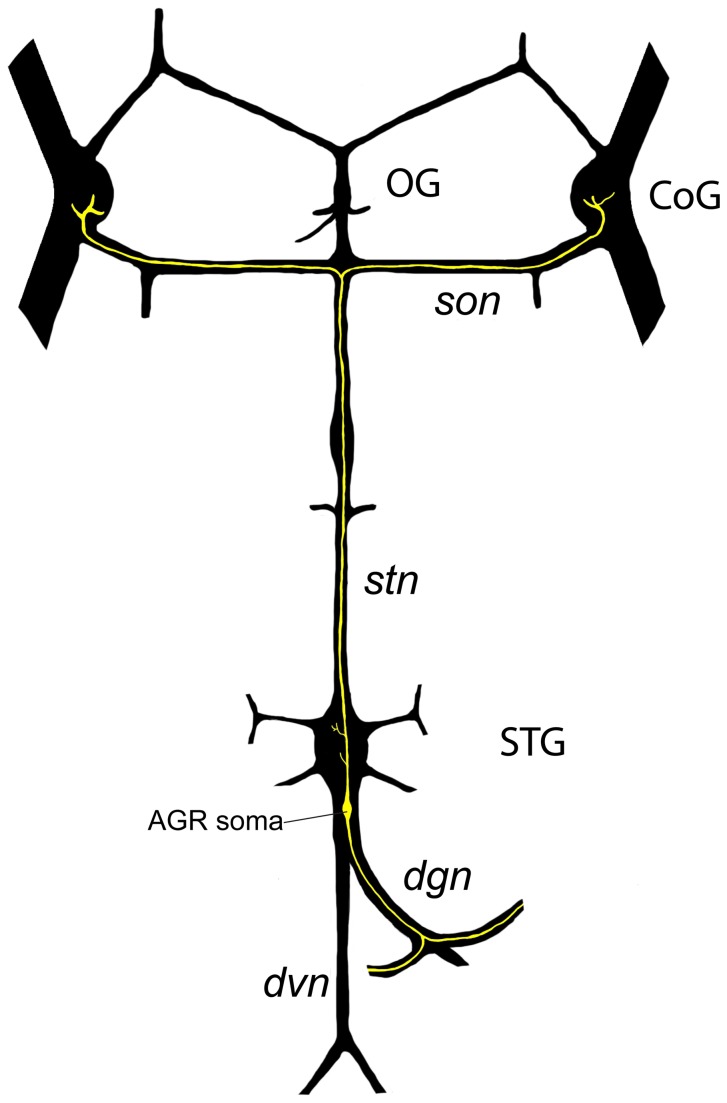
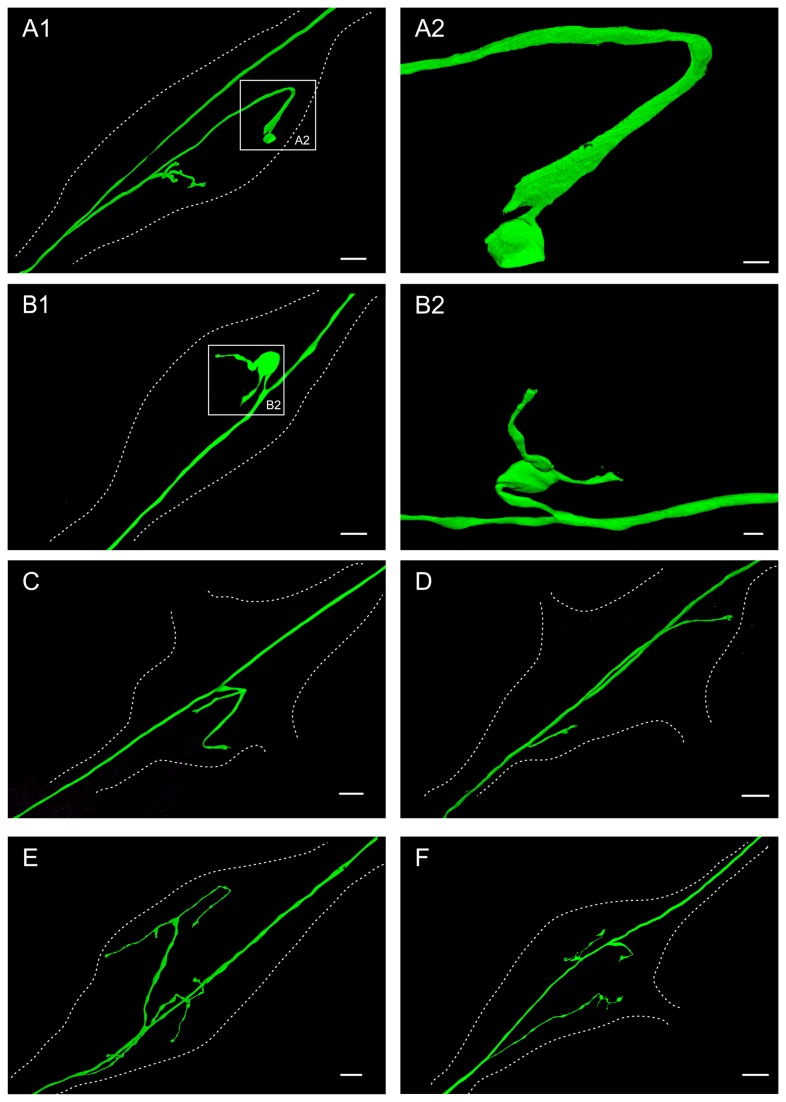
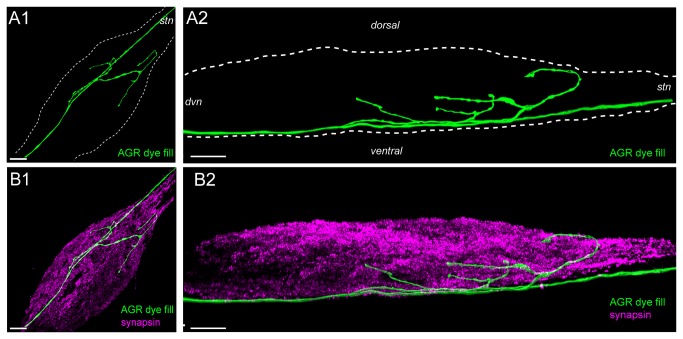
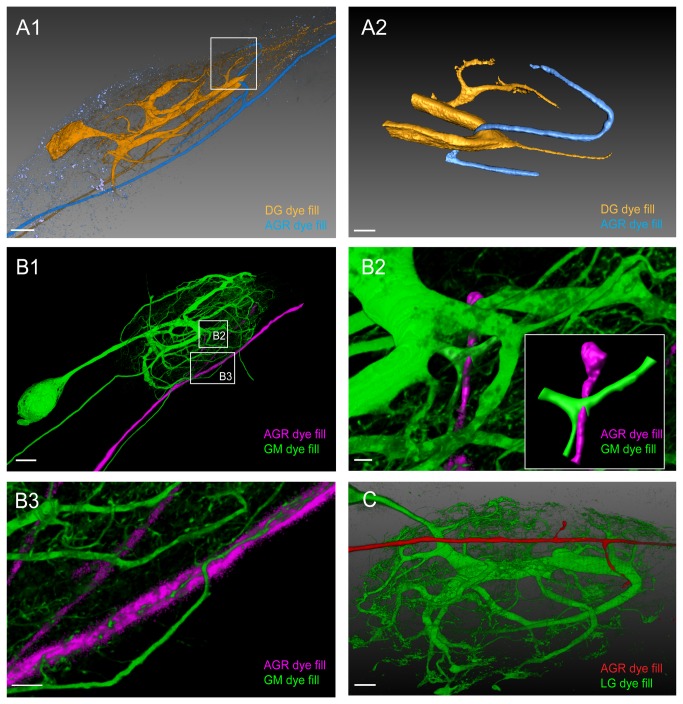
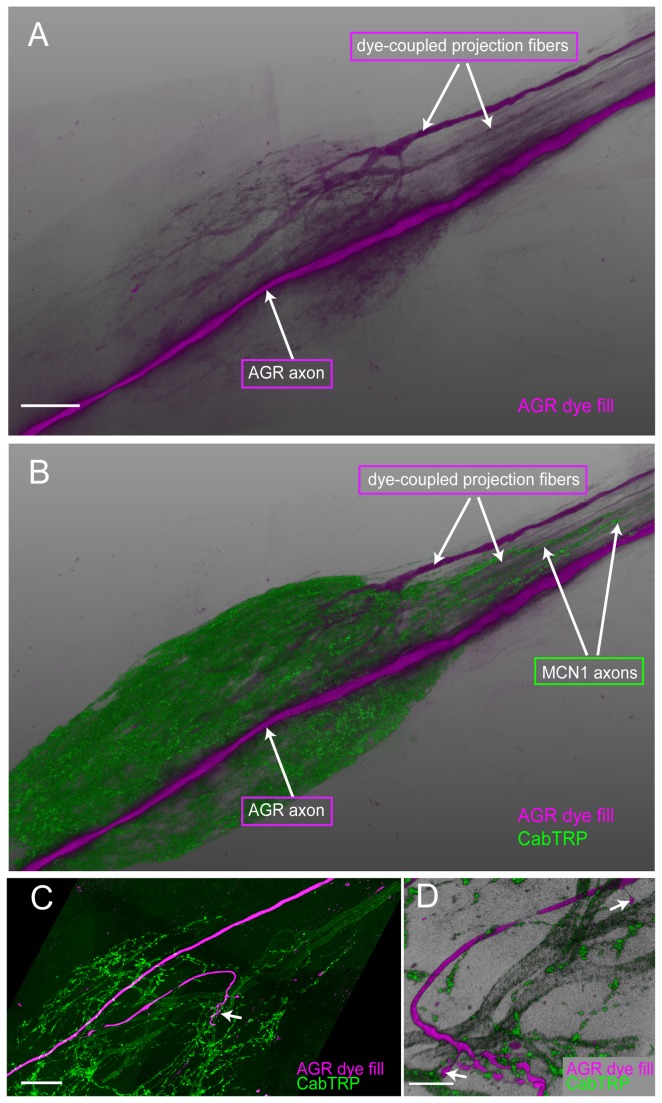
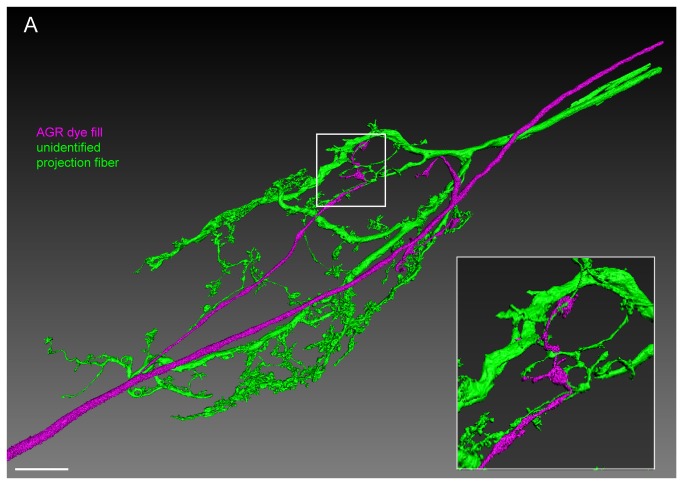
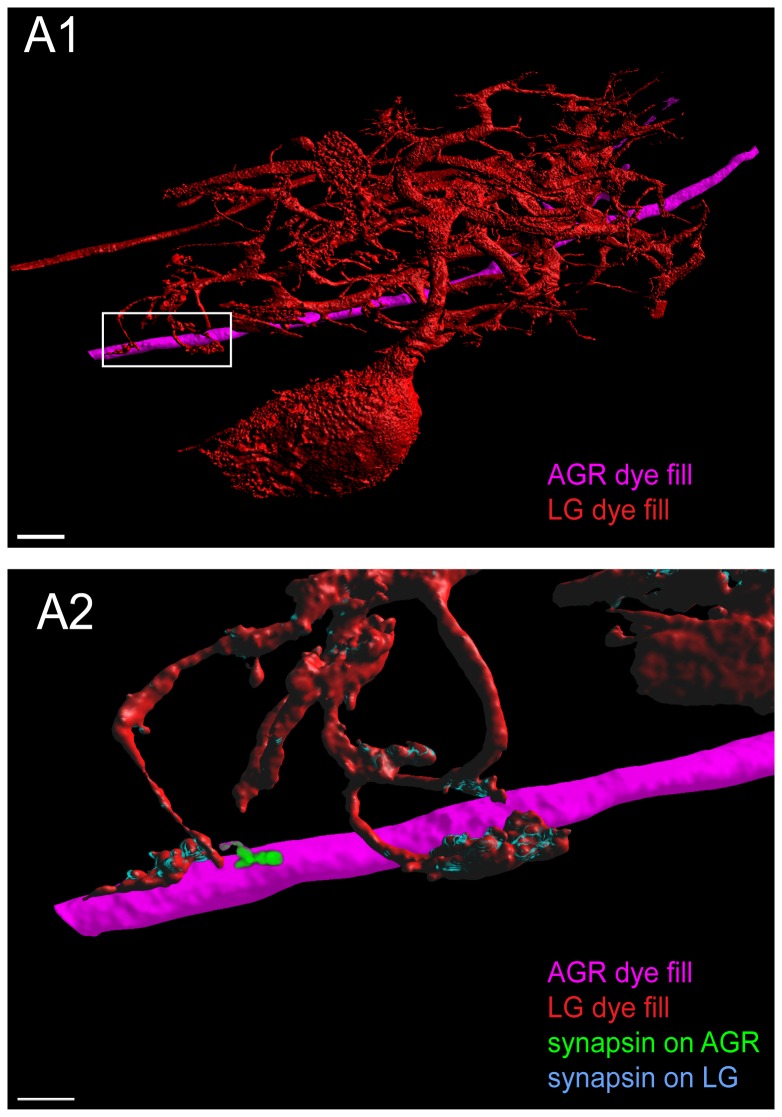
Sensory neurons provide important feedback to pattern-generating motor systems. In the crustacean stomatogastric nervous system (STNS), feedback from the anterior gastric receptor (AGR), a muscle receptor neuron, shapes the activity of motor circuits in the stomatogastric ganglion (STG) via polysynaptic pathways involving anterior ganglia. The AGR soma is located in the dorsal ventricular nerve posterior to the STG and it has been thought that its axon passes through the STG without making contacts. Using high-resolution confocal microscopy with dye-filled neurons, we show here that AGR from the crab Cancer borealis also has local projections within the STG and that these projections form candidate contact sites with STG motor neurons or with descending input fibers from other ganglia. We develop and exploit a new masking method that allows us to potentially separate presynaptic and postsynaptic staining of synaptic markers. The AGR processes in the STG show diversity in shape, number of branches and branching structure. The number of AGR projections in the STG ranges from one to three simple to multiply branched processes. The projections come in close contact with gastric motor neurons and descending neurons and may also be electrically coupled to other neurons of the STNS. Thus, in addition to well described long-loop pathways, it is possible that AGR is involved in integration and pattern regulation directly in the STG.
Conflict of interest statement
Figures

References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
