

Genetic dissection of the polyoxin building block-carbamoylpolyoxamic acid biosynthesis revealing the "pathway redundancy" in metabolic networks
- PMID: 24314013
- PMCID: PMC4029187
- DOI: 10.1186/1475-2859-12-121
Genetic dissection of the polyoxin building block-carbamoylpolyoxamic acid biosynthesis revealing the "pathway redundancy" in metabolic networks
Abstract
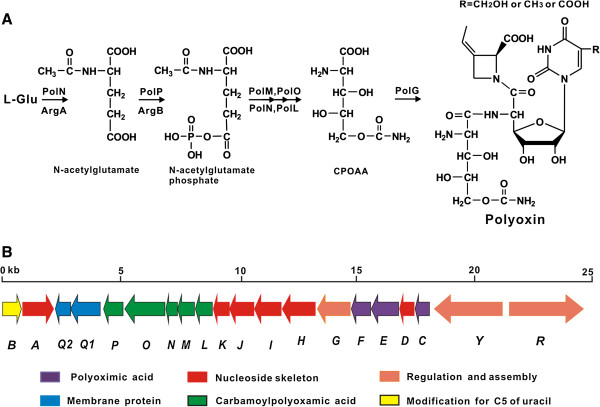
Background: Polyoxin, a peptidyl nucleoside antibiotic, consists of three building blocks including a nucleoside skeleton, polyoximic acid (POIA), and carbamoylpolyoxamic acid (CPOAA), however, little is known about the "pathway redundancy" of the metabolic networks directing the CPOAA biosynthesis in the cell factories of the polyoxin producer.
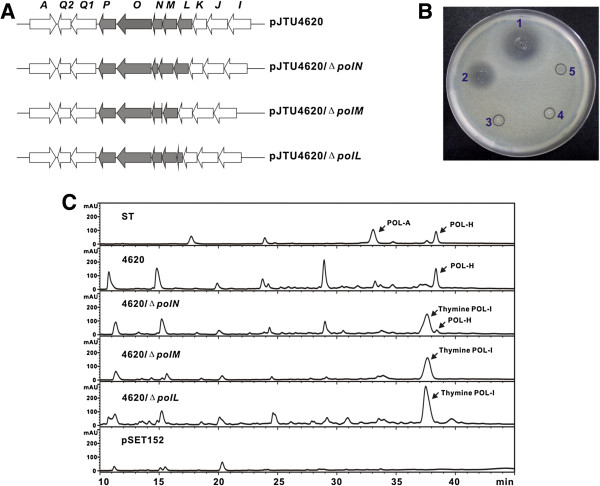
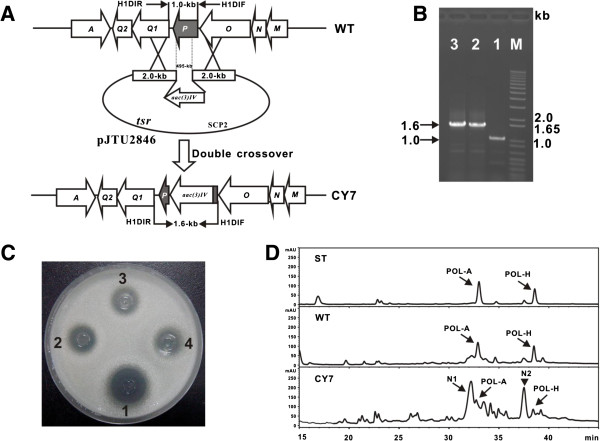
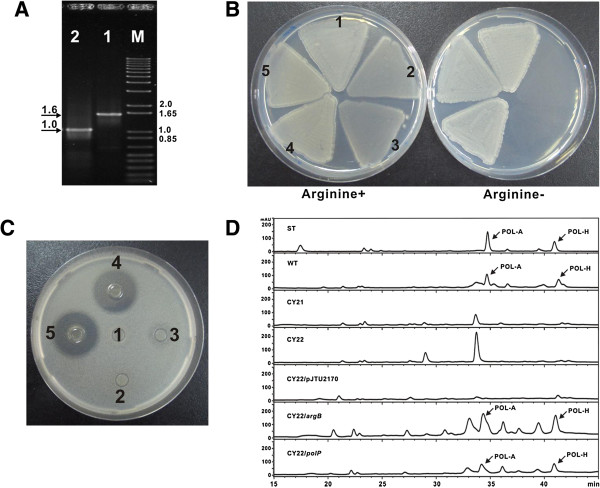
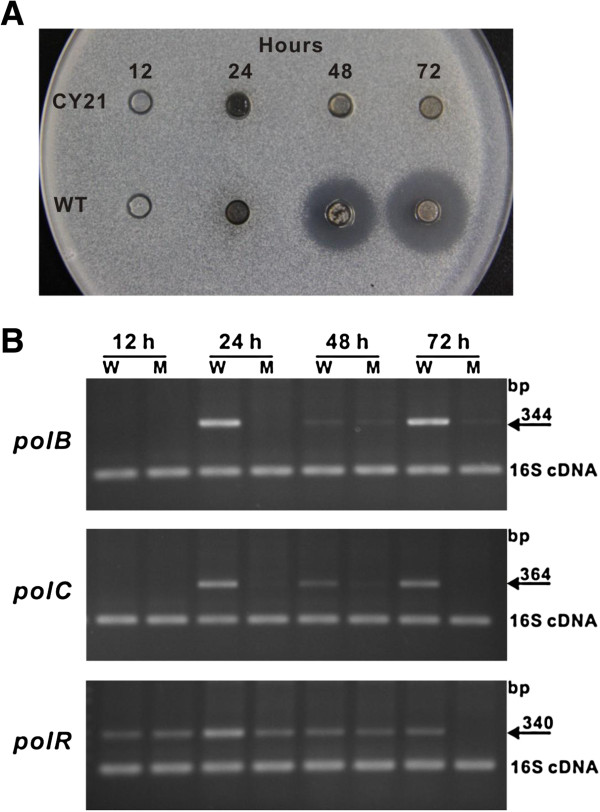
Results: Here we report the genetic characterization of CPOAA biosynthesis with revealing a "pathway redundancy" in metabolic networks. Independent mutation of the four genes (polL-N and polP) directly resulted in the accumulation of polyoxin I, suggesting their positive roles for CPOAA biosynthesis. Moreover, the individual mutant of polN and polP also partially retains polyoxin production, suggesting the existence of the alternative homologs substituting their functional roles.
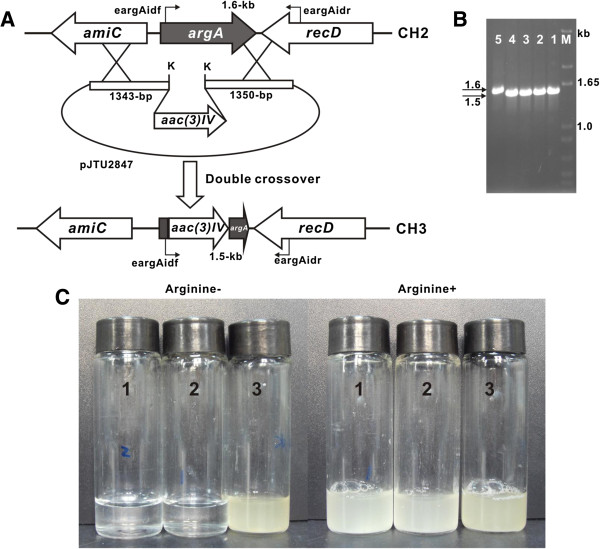
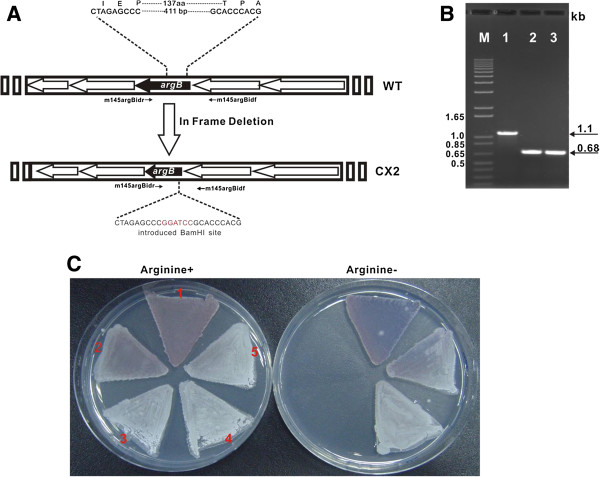
Conclusions: It is unveiled that argA and argB in L-arginine biosynthetic pathway contributed to the "pathway redundancy", more interestingly, argB in S. cacaoi is indispensible for both polyoxin production and L-arginine biosynthesis. These data should provide an example for the research on the "pathway redundancy" in metabolic networks, and lay a solid foundation for targeted enhancement of polyoxin production with synthetic biology strategies.
Figures
References
-
- Suzuki S, Isono K, Nagatsu J, Mizutani T, Kawashima Y, Mizuno T. A new antibiotic, polyoxin A. J Antibiot (Tokyo) 1965;12:131. - PubMed
-
- Isono KN, Kobinata J, Sasaki K, Suzuki S. Studies on polyoxins antifungal antibiotics part V: isolation and characterization of polyoixns C, D, E, F, G. Hand I Agri Biol Chem. 1967;12:190–199. doi: 10.1271/bbb1961.31.190. - DOI
-
- Isono KN, Kawashima J, Suzuki YS. Studies on polyoxins, antifungal antibiotics part I: isolation and characterization of polyoxins A and B. Agr Biol Chem. 1965;12:854.
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
