

Inhibition-induced theta resonance in cortical circuits
- PMID: 24314731
- PMCID: PMC3857586
- DOI: 10.1016/j.neuron.2013.09.033
Inhibition-induced theta resonance in cortical circuits
Abstract
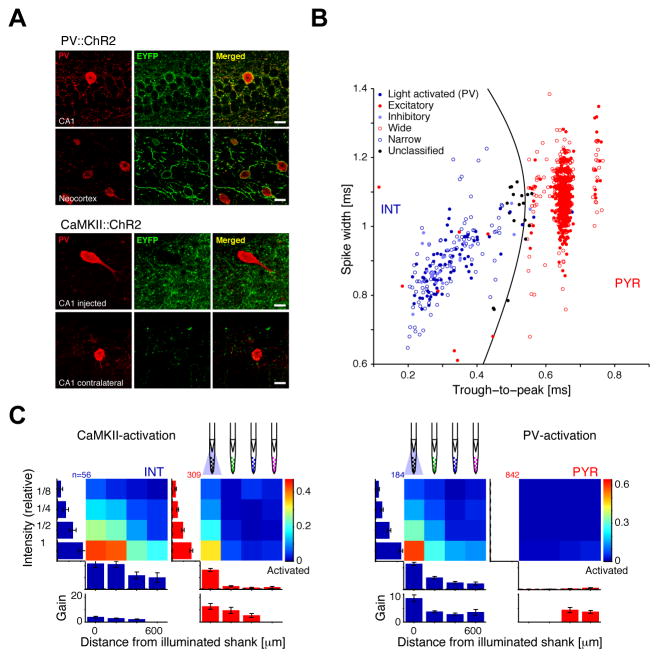
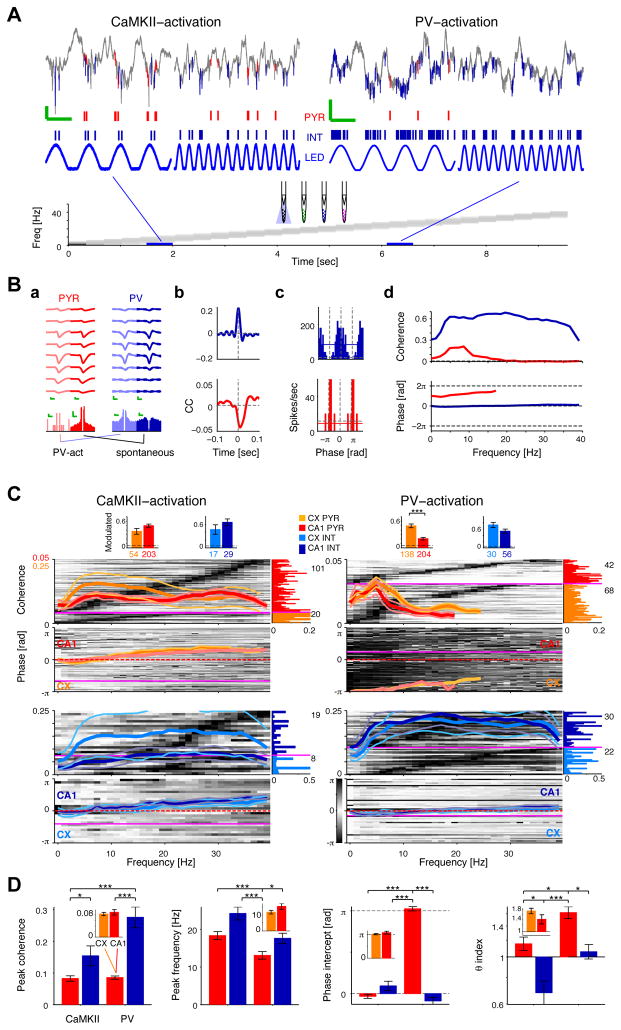
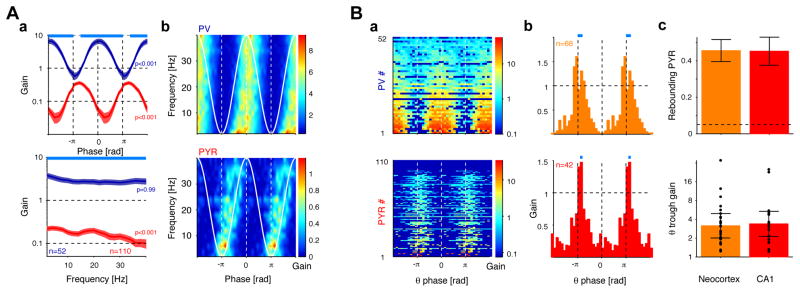
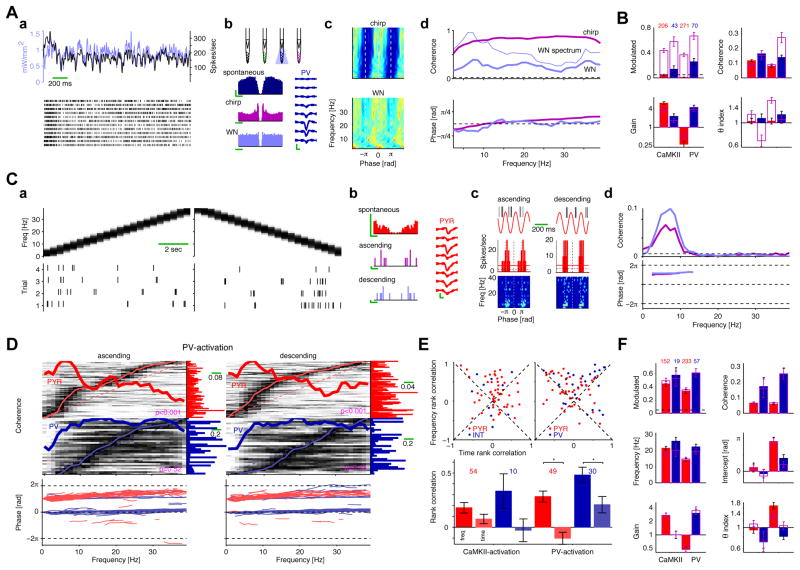
Both circuit and single-cell properties contribute to network rhythms. In vitro, pyramidal cells exhibit theta-band membrane potential (subthreshold) resonance, but whether and how subthreshold resonance translates into spiking resonance in freely behaving animals is unknown. Here, we used optogenetic activation to trigger spiking in pyramidal cells or parvalbumin immunoreactive interneurons (PV) in the hippocampus and neocortex of freely behaving rodents. Individual directly activated pyramidal cells exhibited narrow-band spiking centered on a wide range of frequencies. In contrast, PV photoactivation indirectly induced theta-band-limited, excess postinhibitory spiking in pyramidal cells (resonance). PV-inhibited pyramidal cells and interneurons spiked at PV-inhibition troughs, similar to CA1 cells during spontaneous theta oscillations. Pharmacological blockade of hyperpolarization-activated (I(h)) currents abolished theta resonance. Inhibition-induced theta-band spiking was replicated in a pyramidal cell-interneuron model that included I(h). Thus, PV interneurons mediate pyramidal cell spiking resonance in intact cortical networks, favoring transmission at theta frequency.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
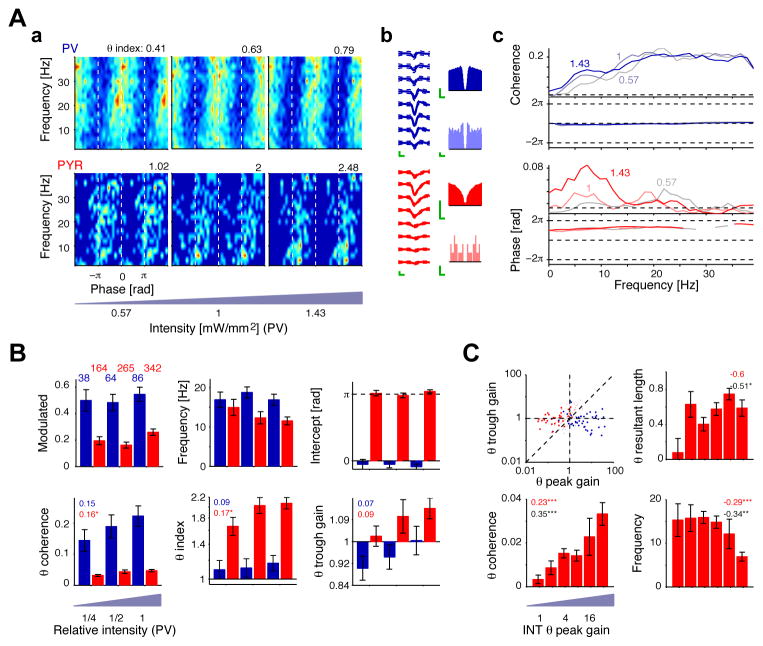
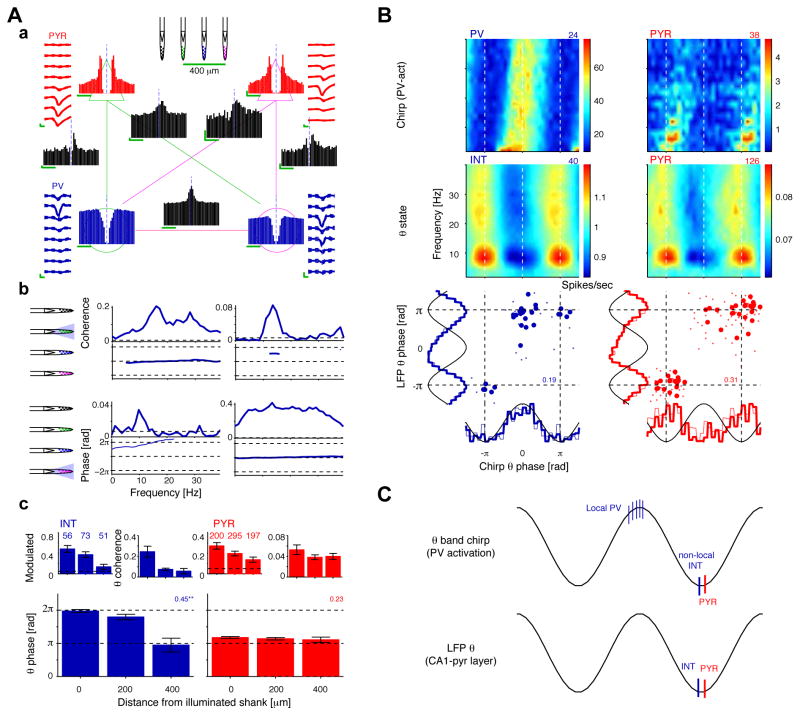
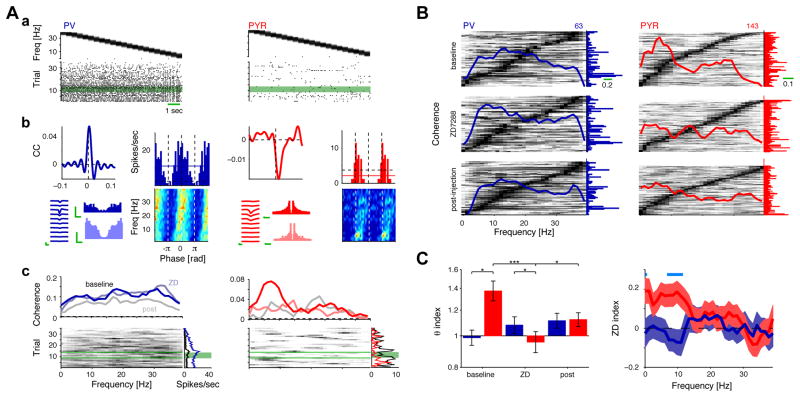
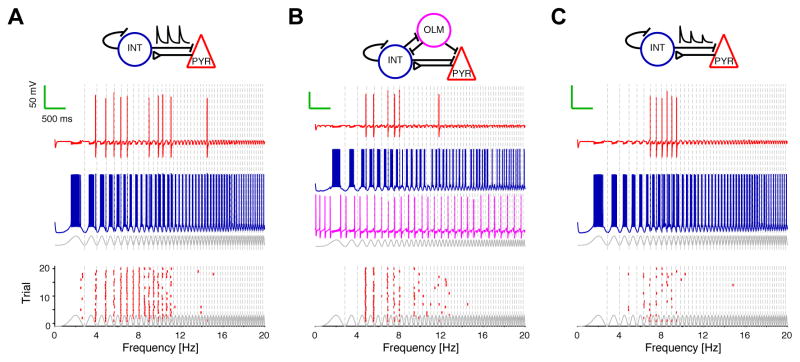
References
-
- Barthó P, Hirase H, Monconduit L, Zugaro M, Harris KD, Buzsáki G. Characterization of neocortical principal cells and interneurons by network interactions and extracellular features. J Neurophysiol. 2004;92:600–608. - PubMed
-
- Boyden ES, Zhang F, Bamberg E, Nagel G, Deisseroth K. Millisecond-timescale, genetically targeted optical control of neural activity. Nat Neurosci. 2005;8:1263–1268. - PubMed
-
- Buzsáki G. Theta oscillations in the hippocampus. Neuron. 2002;33:325–340. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
